Design vs. Descent: A Contest of Predictionsby Casey Luskin Introduction: The famous philosopher of science Karl Popper wrote that all scientific theories must be falsifiable. In order for a theory to be held tentatively and have an empirical basis, Popper argued that it must be subject to falsification and it must be able to be subjected to meaningful tests: In fact, many critics of intelligent design, such as the National Academy of Sciences, have argued that creationist ideas do not qualify as science because they are "not subjectable to meaningful tests, which require predicting plausible results and then checking these results through observation and experimentation3 or "are not science because they are not testable by the methods of science.3" Yet, in the same breath, these critics argue that, "[s]cientists have considered the hypotheses proposed by creation science [previously defined to refer to young earth creationism, old earth creationism, and intelligent design] and rejected them because of a lack of evidence.3" Two questions must now be asked: 1) How can a theory be unfalsifiable, and therefore supposedly unscientific, and yet be falsified by scientific evidence? and 2) Does evolution itself qualify as a scientific theory which can be falsified by scientific evidence? The answer to question 1 in fact is quite simple. A theory which can be said to be falsified by scientific evidence is indeed falsifiable, and by Karl Popper's criteria is a legitimate scientific theory. The National Academy of Sciences, as quoted above, has made a logical fallacy--making two claims which cannot possibly both be true. Indeed, many forms of "creation science" make a variety of predictions which are empirically testable. One example is that young earth creationism predicts that various observable lines of evidence will be found to show that rocks, the oceans, and the solar system are young. Regardless of whether or not such evidence is found, the theory makes scientific predictions which can be tested in the natural world, and thus qualifies the young earth hypothesis as a falsifiable and testable scientific theory. Though different from creation science in very important ways, intelligent design, like any good scientific theory, makes testable predictions. The way that a scientific theory makes predictions is by looking at the mechanism inherent within the theory and asking what sort of results would be expected where that mechanism has been at work. From our observations about how intelligent agents operate when designing, we can understand that intelligent designers act in specific ways when designing that can lead to predictions, as seen in Table 1.
We can then use this understanding of the mechanism of intelligent design to make a few predictions, shown in Table 2:
Each of these predictions can be tested through looking at the scientific data and performing experiments. These robust predictions, based off of what we would expect from the mechanism of design through intelligent agency, refute the common allegation that intelligent design is merely a negative argument against evolution and makes no positive predictions. Intelligent design passes the test of "falsifiability" and therefore meets the qualification of "testability." Before proceeding further, it should be noted that each of these predictions must be evaluated on a case-by-case scenario. Intelligent design theory, like any scientific hypothesis, is meant to be tentative, and invoked only when the evidence warrants.43 However, in contrast to some forms of creation science, intelligent design does not make untestable statements, such as those regarding the identity of the designer. Creation science often explicitly states that "God did this", or "God did that," making claims about the supernatural which seem difficult to test through scientific means. However intelligent design theory makes no such claims about the identity of the designer or the actions of the supernatural and therefore reduces its claims to those which are based upon an empirical understanding of how designers act during intelligent design. In short, it limits its claims and predictions to those which are empirically based, and therefore are testable. Claims from creation science regarding the identity of the designer may be true, however they do not qualify as "scientific claims" (a claim need not be scientific to be true, it only need to be testable to be scientific--some claim which are true may not be empirically testable). However, the second question must still be answered: is Darwinian theory testable and falsifiable? Design versus Descent In The Methodological Equivalence of Design & Descent: Can There Be a Scientific "Theory of Creation"?, philosopher of science Stephen C. Meyer frames this question as "design versus descent," in a competition of which best qualify as strong scientific theories. Meyer uses the term "descent" to refer to all theories of origins which uphold the claim that all-life arose from non-life and is interrelated through ancestry through a purely unbroken naturalistic chain of events (one example is Darwinian evolution). Meyer finds that any argument used against design to disqualify it as a scientific theory can also be used against descent, or some other scientific theory and concludes that a consistent philosophy of science must allow design as a scientific theory, or else allow for the exclusion of other theories currently considered scientific--including descent! Meyer finds that fields studying "design" or "descent" are really historical sciences, such as those which investigate the origins of life, are not able to establish their claims as solidly as many fields of science which test their theories in the present. Most claims from such fields of historical science limit their claims to "inferences" about the past, and can rarely use deductive logic to strongly establish claims through experiment. In other words, it is more difficult to infer the events of the past than to investigate the causes of the present. Meyer concludes that "design" and "descent" are actually epistemological equals: both are equally qualified as bona fide scientific theories:
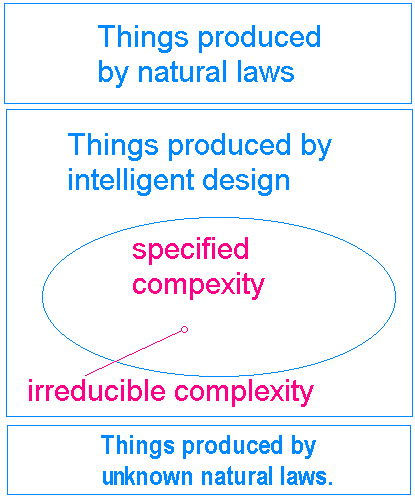
Each of the predictions of design will now be tested against the predictions of descent. While the complexities of these issues might warrant much deeper discussions, the purpose of this document is to outline the evidence as it plays out in the debate over design and descent. For deeper discussions of some of these claims, please see documents linked from appropriate sections. 1. The origin of irreducibly complex systems Darwin himself realized that there were certain types of biological structures which Darwinian evolution simply couldn't create. Darwinian evolution works by a process called the mutation-selection mechanism. Purely random mutations in the DNA create new types of organisms, and those organisms which are better at surviving and reproducing tend to leave more offspring. Over time, populations of organisms change as those which are best suited to the environment get "selected" (i.e. they survive and reproduce better). The catch to all of this is that changes must occur at a very slow pace, one little mutation at a time. Also, biological systems produced by evolution must be functional (i.e. confer some benefit to the organism) at every little step along their evolutionary path. In his book Darwin's Black Box, Michael Behe noted that: 'If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down.' A system which meets Darwin's criterion is one which exhibits irreducible complexity. By irreducible complexity I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning.27 Yet evolutionists do not admit that their theory is falsified by the presence of these unevolvable structures. Instead they turn to explanations of gene duplication and co-optation to build these complex structures.3 While these sorts of explanations still leave the details to the dice and lack strong explanatory power, it should be noted that evolutionists have not allowed their theory to be falsified. As long as there is some protein with some homology to some part in the irreducibly complex structure, evolutionists believe it could have been put their through some combination of gene duplication, co-optation, and micromutation. Thus, evolution makes essentially untestable claims of high improbability to explain the origin of irreducibly complex systems. (See our Irreducible Complexity Page and Nonfunctional Intermediates Page for more details on these issues.) In intelligent design, irreducible complexity is merely a special case of "specified complexity" (see Figure 1).
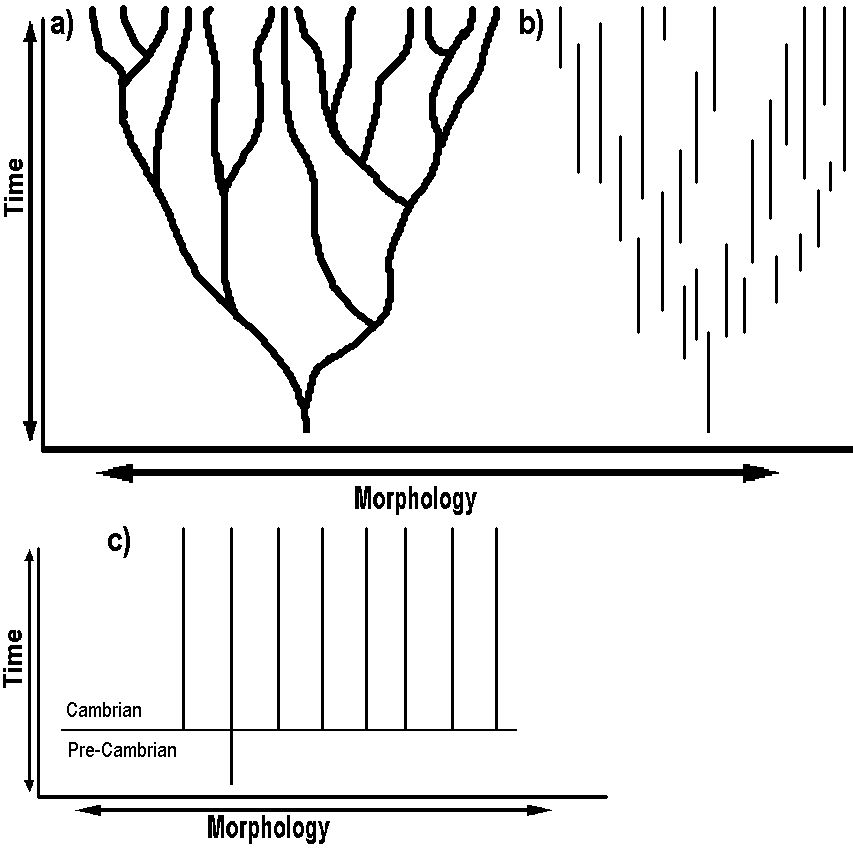
One hot area of research in biology relevant to looking for "specified complexity" is investigating protein specificity. Many new articles have been published in recent years, and many new research groups are beginning to ask questions about how specific biological parts must be in order to function. Three mainstream articles include: Each article concludes that proteins and thus DNA are highly specified in the function they perform. This sort of research is on its way to detecting specified complexity, and affirming a design inference, and is an active area of research for intelligent design theorists. As far as the flagellum goes, it almost looks as if the design inference is on its way to being made, as one biologist said in a major mainstream biology journal, Cell, "[m]ore so than other motors, the flagellum resembles a machine designed by a human."35 2. The fossil record and transitional forms Darwin himself realized that his theory predicted that fossils indicating transitionss from one form to another should be found in the fossil record. For this reason, Darwin asked the question:
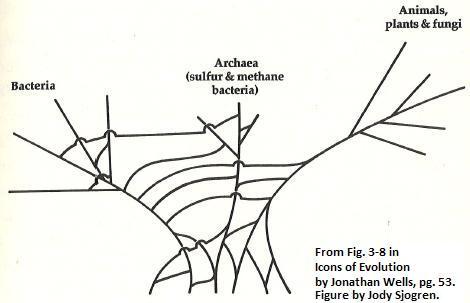
3. Darwinian trees and molecular evolution "Family Trees" (called "phylogenetic trees") based off of DNA sequences in genes should make nice neat "Darwinian Trees" if common ancestry is true. However, it is well recognized in the field of classification (systematics) that very often phylogenetic trees based upon one gene or protein sequence, will lead to one tree, while a tree based upon some other biomolecule will look quite different. Molecular biologist W. Ford Doolittle wrote, "[m]olecular phylogenists will have failed to find the 'true tree,' not because their methods are inadequate or because they have chosen the wrong genes, but because the history of life cannot properly be represented as a tree."7 Yet, evolution predicts that molecular data should allow a phylogenetic "tree of life" to be reconstructed. Descent, apparently, has been falsified. As Doolittle indicates, from the base of the tree of life, it is not "tree-like." In the "bush" below (Figure 3), it is impossible to reconstruct such trees, as the observed distribution of characters create something which looks more like a tangled thicket or a bush. The three major "domains" of life--Bacteria, Archaea, and Eukarya have a distribution of characteristics which does not allow a tree to be constructed to describe their alleged ancestral relationships. This is due to a character distribution which is not what one would predict if they inherited their genes through common ancestry: 1. Ribosomal genes in archaea are similar to those found in eukaryotes 2. Morphology of archea is much more similar to bacteria. 3. Eukaryote "operational" genes (those involved with amino acid synthesis and metabolism) seem to be most similar to those found in bacteria.7, 11 4. Eukaryote "informational" genes (those involved with cell replication) seem to be most similar to those found in Archaea.7, 11
Though some have envisioned the origin of Eukaryotes as a symbiotic fusion of members of archaea with members of bacteria, this simple "symbiosis hypothesis" has been recently challenged after it was found that eukaryotes contain over 300 proteins which do not bear any resemblance to any proteins in their supposed bacteria and archaea ancestors. The researchers suggest that a hypothetical pre-cursor cell type--the chronocyte--merged with members of both the archea and bacteria domains to provide these extra genes, making the an evolutionary symbiotic origin of eukaryotes a much more complicated and much less elegant theory.8 The tree of life, at its base, is nothing of the sort, and predictions from descent have failed. Many defenders of descent have argued that gene sharing, differing rates of evolution, or even convergent evolution among microorganisms is perhaps a plausible explanation for the lack of consensus among trees. But regardless, a prediction of the descent hypothesis has again failed, and we are left with an "epicycle". Descent-based theories become so plastic that they can potentially explain any dataset, and there is no way to falsify the theory as it loses explanatory value. Just as the case with punctuated equilibrium, descent becomes unfalsifiable with respect to molecular phylogenetic trees. The father of molecular systematics Carl Woese found that conflicts in phylogenies are present not only at the base of the tree, saying, "[p]hylogenetic incongruities [conflicts] can be seen everywhere in the universal tree, form its root to the major branchings within and among the various taxa to the makeup of the primary groupings themselves."21 For example, de Jong noted that,"the wealth of competing morphological, as well as molecular proposals [of] the prevailing phylogenies of the mammalian orders would reduce [the mammalian tree] to an unresolved bush, the only consistent clade probably being the grouping of elephants and sea cows.12" Another recent study entitled "Widespread horizontal transfer of mitochondrial genes in flowering plants" invoked the gene-swapping mechanism to explain the fact that some plant mitochondrial genes are distributed in patterns strongly at odds with expectations from common descent.44 At least one study would argue that horizontal gene transfer isn't even possible in advanced Eukaryotes like plants.46 Cao et al. found that molecular-based phylogenies conflicted sharply with previously established phylogenies of major mammal groups, such as ferungulates, rhodents, and primates.22 Some studies have tried to analyze the general relationships between animals and vertebrate groups through molecular data. One study analyzed molecular data from 10 different vertebrates and found that using different mitochondrial genes, twenty different disagreeing phylogenetic trees were produced, which differed at both recent and ancient divergence points.23 Brown and Naylor24 found that trees based off of entire mitochondrial DNA genomes for 19 multicellular (metazoan) organsism did not match the previously accepted phylogeny for chordates, or within chordates, for vertebrates. Finally, a study which compared many proteins in humans, nematodes, arthropods, and yeast found that 2 starkly different trees were produced, depending on which genes were used.25 This pattern of different genes yielding very different phylogenetic trees is very common in the scientific literature, and shows that molecular data fail to give a consistent picture of the alleged common descent ancestry of organisms. It has also been recognized that various genes distributed in organisms often do not match predictions of Darwinism. Schwabe and Warr found that proteins such as relaxin, insulins, adrenocorticotropic hormone, somatostatin, histocompatability antigens, neural glycoproteins and microglobulin are distributed in both animal and non-animal groups in ways which differ markedly from predictions of common descent.13 Another prediction of descent is that molecular data should verify phylogenetic trees based upon "macro-morphology" (larger structural features of biological organisms) in both living and fossil organisms. However, molecules and morphology often strongly disagree, and as one researcher put it, this causes 'evolution wars': Systematist Colin Patterson also did a review of congruence between trees generated by morphology and trees created by molecular data and found that "congruence between molecular phylogenies is as elusive as it is in morphology and as it is between molecules and morphology."14 His study concluded that after reviewing trees based off of molecular and morphological data, their hopes of verifying morphologically derived trees were "dampened,"14 disconfirming their expectations from descent-based hypotheses. Some have hoped that over time, as more data is accumulated by finding more fossils and determining more DNA sequences, that apparent contradictions between the molecular and morphologically-based trees would be resolved. However, a recent study reported that, "our ability to reconstruct accurately the tree of life may not have improved significantly over the last 100 years" and that, "[d]espite increasing methodological sophistication, phylogenies derived from morphology, and those inferred from molecules, are not always converging on a consensus."26 Despite 100 years of intense and disconfirming research, the study attributed the contradictions to a lack of data. Molecular-based trees are often at odds with trees created by the fossil record, or from structural characteristics of organisms. This is a serious problem for "descent," because it shows that evidence for common ancestry (the idea that all life-forms are related through ancestry) is not supported or flatly contradicted by molecular data. Proponents of descent invent a number of "ad hoc" hypotheses to explain away the inconsistencies (gene sharing, changing rates of evolution, convergent evolution, lack of data) however descent has now been placed in an unfalsifiable position also with respect to molecular data. 4. Allegedly "Junk DNA" or "non-functional DNA." Ever since scientists began to understand the working of genetics, it became apparent that only about 3% of our DNA is composed of genes that code for proteins. Many evolutionists have claimed that the rest of the DNA is junk-DNA," an evolutionary relic of ancient discarded genes. Not only do they claim it is a fulfilled prediction of evolution, but it is seen as a non-functional product that an intelligent designer wouldnt create. Yet, in recent years, more and more types of junk-DNA have been found to have function. If this trend continues, perhaps eventually most DNA will be found to have function (though probably not all as some information loss has undoubtedly occurred whether it was originally designed or not). Consider the following examples: An extensive discussion of examples of functionality of previously alleged "Junk DNA" can be found at When "Junk" DNA Isn't Junk (http://www.godandscience.org/evolution/junkdna.html). It should be noted that various types of DNA persist whose functions are not yet known. However, given the current trend that as we increase understanding of genetics, many many more types of previously thought "junk-DNA" is found to be functional, one might expect that in time, many, if not all types of DNA might be found to have function. In the end, perhaps it will turn out that expectations from evolutionary theory that the genome should be full of relic "junk-DNA" might have actually hindered and slowed our investigation and understanding of many types of DNA. It should also be noted that intelligent design theory only predicts original functionality, and allows for loss of function, where something was originally designed functional, but perhaps function was lost due to various environmental factors. This could be seen as analagous to leaving brand new car out on a mountaintop. 500 years later, natural forces leave parts of it non-functional, although it was designed as intially functional. It is also possible that a designer might have, in some instances, pre-programmed diversity as "dormant genes" which might have been unexpressed in the past, or expressed in the past but unexpressed in the present. This "pre-programmed diversity" could account for some redundant, but functional-gene-like components of "junk-DNA" whose functions are currently unknown, as is the case with some pseudogenes. Thus "functionless DNA" might be designed to be only "temporarily functionless" and not the permanently functionless evolutionary relic that evolutionists mistake them to be. Expectations and predictions of design seem to be yielding much more fruit in this area of research than are predictions of descent via neo-Darwinian processes. Evolution: A Theory on Life Support? In his book, "The Structure of Scientific Revolutions", historian and philosopher of science Thomas Kuhn argues that a scientists will add "ad hoc" explanations in order to cover up inconsistencies between the data and the current paradigm (a paradigm is a dominant theory dictating the research and interpretation of data in a field of science). Kuhn recognized that in the face of disconfirming evidence, scientists would much rather "explain away" the inconsistent data through some "auxiliary hypothesis" than to simply disregard the paradigm. For example, before Copernicus developed the heliocentric model of our solar system, most scientists believed that the planets and the sun revolved around the earth. Yet many astronomers of the time noticed that planets sometimes exhibited "retrograde motion" where they would temporarily move backwards in circular paths, unpredicted by the geocentric theory. This, according to geocentrists, was the result of "epicycles" where planets not only revolved around the earth, but also around a central point along their orbital path. An epicycle was therefore an additional explanation used to resolve inconsistencies between the scientific data and predictions from the now-falsified geocentric model of the solar system. Had these astronomers had a different sun-centered theory of the solar system, they would have quickly realized that these "retrograde epicycles" were merely the result of the earth's annual orbit around the sun. In the same way, proponents of naturalistic theories of descent now develop "epicycle-like" explanations to resolve apparently falsifying data. While descent hypotheses could potentially be falsified by this data, these "auxiliary hypotheses" (i.e. punctuated equilibrium, miraculous genetic duplications and co-optations, lateral gene transfer, hopeful lack of data) do serve to preserve the original theory of descent. However, in the process, they force the primary claims of common descent, namely the fact that all organisms are related through ancestry and evolved by mutation and selection, into an unfalsifiable position. In his book, Kuhn notes that a theory in "crisis" will develop such auxiliary hypotheses to save the core tenets of the theory. As seen in the geocentrism example, false hypotheses can survive for hundreds of years before enough "quirks" in the data develop to force some scientists to look elsewhere. However Kuhn observes that scientists will generally not consider abandoning a paradigm or important theory until they are able to replace it with a new paradigm which can better explain the "quirks" of the old paradigm, "...a scientific theory is declared invalid only if an alternative candidate is available to take its place."28 Since the time of Darwin, evolutionists have known about the weaknesses of the theory of evolution and descent with modification.41 Yet, they have not abandoned it simply because they say, "it's the best theory we've got". Descent, through these "ad hoc" hypotheses, has been forced into an unfalsifiable position, lest it be falsified. Today, biologists have a new and growing theory of life's origin: intelligent design. As intelligent design theory matures and develops as a scientific theory, evolution may not be the "best" any longer, and the "design hypothesis" may once again be considered as a viable explanation for the origins of lifeforms on earth. Many have argued that design isn't a potent explanation because it can explain potentially anything. However, if more and more examples of unevolvable high complexity are found in nature, and more and more forms seem to turn up in the fossil record without precursors, and if genes seem to be distributed in a manner where a designer re-used common parts in different organisms, then intelligent design can indeed better explain the data:
Evolutionists currently resolve the discrepancies between its predictions and the data through "ad hoc" hypotheses. However, if evolution cannot explain the data over time, then there is nothing wrong with appealing to a theory which can. Thus, it can be seen from this document that intelligent design theory makes empirically-based predictions, which are matched in the data. Until the time that biologists are ready to admit design into their picture, the quirks and counter-evidences to evolution, and the auxiliary hypotheses of descent-based theories may increase, and the data may become increasingly inconsistent with evolution. Descent has failed each of these tests, yet it has been retained by scientists as a valid scientific description of the history and origins of life on earth. Hopefully, at some point, science will consider design. References Cited: 1. Popper, Karl, Conjectures and Refutations (New York: Basic Books, 1963), p. 257. 2. See essays by Larry Lauden in But Is It Science? Edited by Michael Ruse. Buffalo, N.Y.: Prometheus Books, 1988. 3. Science and Creationism A view from the National Academy of Sciences (Second Edition), 1999, National Academy Press. 4. The Methodological Equivalence of Design & Descent: Can There Be a Scientific "Theory of Creation"? by Stephen C. Meyer, posted online at http://www.arn.org/docs/meyer/sm_methodological.htm. 5. Charles Darwin, Origin of Species. 6. Stephen Jay Gould, 'Is a new and general theory of evolution emerging?' Paleobiology, vol 6(1), January 1980, pg 127. 7. Doolittle, W. F. Phylogenetic Classification and the Universal Tree. Science, Vol 284:2124-2128 (June 25, 1999). 8. The origin of the eukaryotic cell: A genomic investigation in Proc. Nat. Acad. Sci. USA Vol. 99, Issue 3, pg. 1420-1425, February 5, 2002 by Hyman Hartman and Alexei Fedorov. 9. The Design Inference by William Dembski (Cambridge University Press, 1998) 10. Dehaan, R. F. and Wiester, J. L., "The Fossil Record and Intelligent Design" in Signs of Intelligence, edited by William Dembski and James M. Kushiner (pg. 145-156). 11. Rivera M, Jain R, Moore J, Lake J. Genomic Evidence for Two Functionally Distinct Gene Classes. Proc. Nat. Acad. Sci. USA, Vol. 95, pg. 6239-6244, June, 1998. 12. De Jong, W. W. Molecules remodel the mammalian tree. Tree Vol 13, No 7, pg. 270-274 (July 7, 1998). 13. Schwabe, C., Warr, G. W., "A polyphyletic View of Evolution: The Genetic Potential Hypothesis." Perspectives in Biology and Medicine, 27, 3, Spring 1984 pg. 465-484. See also "On the validity of molecular evolution" by Christian Schwabe (TIBS 11 - July 1986 pg. 280-283). 14. Patterson, C., Williams, D. M., Humphries, C. J. "Congruence Between Molecular and Morphological Phylogenies". Annual Review Ecology Systematics 24:153-88 (1993). 15. "Bones, Molecules or Both" by Trisha Gura in Nature vol 406, 230 - 233 (2000). 16. Milinkovitch , M C., "Molecular phylogeny of cetaceans prompts revision of morphological transformations". Trends in Ecology and Evolution 10: 328-334 (August 1995). 17. Hasegawa, M., Adachi, J., Milinkovitch, M. C. "Novel Phylogeny of Whales Supported by Total Molecular Evidence," Jour. of Mol. Evol. 44: S117-S120 (Supplement 1, 1997). 18. Normile, Dennis, "New Views of the Origins of Mammals, (a report on the International Symposium on the Origin of Mammalian Orders, July 21-15, 1998) Science 281:773-774. 19. Novacek, M. J., "Mammalian phylogeny: shaking the tree," March 12, 1992. Nature 356:121-125. 20. Hedges, S. B., Poling, L. L., "A Molecular Phylogeny of Reptiles," Science 238:998-1001 (February 12, 1999). 21. Woese C. The Universal Ancestor. Proc. Nat. Acad. Sci. USA, June 1998, 95:6854-9859. 22 Cao Y, Janke A, Waddell P, Westerman M, Takenaka O, Murata S, Okada N, Paabo S, Hasegawa M. Conflict Among Individual Mitochondrial Proteins in Resolving the Phylogeny of Eutherian Orders. J. Mol Evol, 47:307-22 (1998). 23 Cummings M, Otto S, Wakeley Biolical Bulletin, 196:345-50 (June 1999). 24. Gavin N, Brown W. Amphioxus Mitochondrial DNA, Chordate Phylogeny, and the Limits of Inference Based on Comparisons of Sequences. Syst. Biol. 1998;47(1):61-76. 25. Mushegian A, Garey J, Martin J, Liu L. Large-Scale Taxonomic Profiling of Eukaryotic Model Organisms: A Comparison of Orthologous Proteins Encoded by the Human, Fly, Nematode, and Yeast Genomes. Genome Research 1998;8:590-8. 26. Wills, M. A., "The tree of life and the rock of ages: are we getting better at estimating phylogeny" in BioEssays 24:203-207 (2002) reporting on the findings of Benton, M. J., "Finding the tree of life: mathing phylogenetic trees to the fossil record through the 20th century" in Proc. R. Soc. Lond. B. 268:2123-2130 (2001). 27. Behe, M.J., Darwin's Black Box, pg. 39 (Free Press, 1996). 28. Kuhn, T. S. (1970) The Structure of Scientific Revolutions, 2nd ed, University of Chicago Press, Chicago. 29. For a brief discussion of this matter, see Wells, J., Icons of Evolution pg. 60 (Regnery 2000) or Icons Still Standing by Casey Luskin in the "Homology in Vertebrate Limbs" subsection. 30. This idea has been put forth by a number of design theorists. For one example, see biologist Jonathan Wells' statements as reported in this Jonathan Wells Lecture Report from Western Michigan University or this e-mail report from scientist Phil Skell. 31. DNA and Other Designs by Stephen C. Meyer. 32. No Free Lunch by William Dembski (2001). 33. The Origin of Species by Charles Darwin 34. Hickman, C.P. [Professor Emeritus of Biology at Washington and Lee University in Lexington], L.S. Roberts [Professor Emeritus of Biology at Texas Tech University], and A. Larson. 2000. Animal Diversity. McGraw Hill, NY. 429pp.; (p. 23) 35. DeRosier, D. J., Cell, 93:17 (1998). 36. Fedorov, A., Merican, A.F., and Gilbert, W., "Large-scale comparison of intron positions among animal, plant, and fungal genes," in Proc. Nat. Acad. Sci. USA, 99:16128-16133 (2002). Click here for the abstract. 37. "Initial sequencing and comparative analysis of the mouse genome" (many co-authors), Nature, 420:520-562 (2002). 38. Hakimi, M.S. et. al., "A chromatin remodeling complex that loads cohesin onto human chromosomes," Nature, 418:994-998 (2002). 39. Beaton, M.J. and T. Cavalier-Smith, "Eukaryotic non-coding DNA is functional: evidence from the differential scaling of cryptomonal genomes," Proc. R. Soc. Lond. B. 266:2053-2059 (1999). 40. Morrish, T. A., et. al., "DNA repair mediated by endonuclease-independent LINE-1 retrotransposition," Nature Genetics, 31(2):159-165 (June 2002). 41. This claim is established because Darwin himself recognized various problems with his hypothesis of descent with modification, and discussed them in Origin of the Species. Thus, evolutionists were put on notice of these problems by Darwin at the time he wrote Origin and have effectively known of their existence since Darwin's time. Some of the problems Darwin recognized, which are also discussed in this article, include: 1. Biological organs or systems which cannot be formed through the mutation-selection mechanism. Darwin recognized this problem when he wrote: "If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down." 2. Lack of transitional forms, recognized by Darwin when he wrote: '"... The number of intermediate varieties, which have formerly existed on the earth, (must) be truly enormous. Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory." 42. Nature 423:91-96 (May 1, 2003)) see also: "Researchers in Japan and UCSD Discover Novel Role For Pseudogenes". 43. There may be many cases where the predictions of intelligent design theory are met. However, there may be others where these predictions are NOT met. Intelligent design theory is meant to be only one possible explanation for the origin of any given biological feature. There may be other possible explanations, such as evolution, for any given biological feature. So, just because some prediction of intelligent design theory are met in one case do not mean that all features are therefore the result of intelligent design--scientific theories are only invoked when warranted by the evidence, and if other cases exist where the predictions of intelligent design are not meant, then intelligent design is, scientifically speaking, not the proper explanation. The converse is also true: just because the predictions of design are NOT met in some cases does not mean that intelligent design is falsified for ALL cases. In scientific reasoning, predictions need to be evaluated and explanations derived on a case-by-case basis, as the evidence warrants. 44. Bergthorsson, U., Adams, K., Thomason, B., Palmer, J. D., "Widespread horizontal transfer of mitochondrial genes in flowering plants", Nature 424, 197 - 201 (July 10,2003). 45. See Lev-Maor, G. et al. The birth of an alternatively spliced exon: 3 splice-site selection in Alu exons. Science 300, 1288-1291 (May 23, 2003); W. Makalowski. Not junk after all. Science 300, 1246-1247 (May 23, 2003); and Junk DNA Creates Novel Proteins from the Genome News Network. 46. "Horizontal gene transfer: A critical view," C. G. Kurland, B. Canback, and Otto G. Berg, PNAS, August 19, 2003, vol. 100, no. 17, pg. 9658-9662 (see "http://www.pnas.org/cgi/content/abstract/100/17/9658"). |
Related Articles: |