By Casey LuskinNon-functionality and Irreducible Complexity:
In the
Origin of the Species, Charles Darwin said,
"If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down."1
In evolution, natural selection only preserves those structures which confer some advantage for the organism. If a structure isn't functional, then it confers no advantage, is a waste of the organism's resources, and will be selected out. Darwin says that there may exist structures for which functional intermediate stages would be impossible, i.e. the intermediates would not function. This is essentially the same challenge of
irreducibly complex structures, where intermediate structures wouldn't be functional. Biologist Michael Behe explains:
"A system which meets Darwin's criterion [listed in the above quote] is one which exhibits irreducible complexity. By irreducible complexity I mean a single system which is composed of several interacting parts that contribute to the basic function, and where the removal of any one of the parts causes the system to effectively cease functioning. An irreducibly complex system cannot be produced gradually by slight, successive modifications of a precursor system, since any precursor to an irreducibly complex system is by definition nonfunctional. Since natural selection requires a function to select, an irreducibly complex biological system, if there is such a thing, would have to arise as an integrated unit for natural selection to have anything to act on. It is almost universally conceded that such a sudden event would be irreconcilable with the gradualism Darwin envisioned."4
In the quote above, Behe notes that there is a fundamental quality of any irreducibly complex system in that, "any precursor to an irreducibly complex system that is missing a part is by definition nonfunctional.
4 Behe elaborates upon this definition saying
"An irreducibly complex evolutionary pathway is one that contains one or more unselected steps (that is, one or more necessary-but-unselected mutations). The degree of irreducible complexity is the number of unselected steps in the pathway."11
More than Just Behe?
Behe, who also compares the problem of functional intermediates to a "groundhog trying to cross a thousand lane highway,"
9 is not alone in his sentiments. Many biologists the problem of non-functional intermediates to be a formidable challenge to Darwin's theory.
Soren Lovtrup, professional biologist in Sweden, said "...the reasons for rejecting Darwin's proposal were many, but first of all that many innovations cannot possibly come into existence through accumulation of many small steps, and even if they can, natural selection cannot accomplish it, because incipient and intermediate stages are not advantageous."
2 Well known evolutionist vertebrate paleontologist Robert Carroll asked if the gradual processes of microevolution can evolve complex structures:
"Can changes in
individual characters, such as the relative frequency of genes for light and dark wing color in moths adapting to industrial pollution, simply be multiplied over time to account for the origin
of moths and butterflies within insects, the origin of insects from primitive arthropods, or the origin of arthropods from among primitive multicellular organisms? How can we explain the
gradual evolution of entirely new structures, like the wings of bats, birds, and butterflies, when the function of a partially evolved wing is almost impossible to conceive?"10
To overcome the problems of non-functional intermediates, some biologists have proposed "macromutations" or "saltations" which would produce radically different organisms. Though Stephen Jay Gould is not a proponent of this theory, he noted that, "the absence of fossil evidence for intermediary stages between major transitions in organic design, indeed our inability, even in our imagination, to construct functional intermediates in many cases, has been a persistent and nagging problem for gradualistic accounts of evolution."
8 Those who proposed that rare macromutations would produce "hopeful monsters", some of which might actually have some great advantage, have not been received well by biologists. Paleobiologists Douglas Erwin and James Valentine explain why:
"Viable mutations with major morphological or physiological effects are exceedingly rare and usually infertile; the chance of two identical rare mutant individuals arising in sufficient propinquity to produce offspring seems too small to consider as a significant evolutionary event. These problems of viable "hopeful monsters" ... render these explanations untenable."3
Erwin and Valentine said this in regards to the origin of the major body plans of life--the phyla--and some marine classes, however, others have found other unevolvable structures. Turkish evolutionist Engin Korur says, "The common trait of the eyes and the wings is that they can only function if they are fully developed. In other words, a halfway-developed eye cannot see; a bird with half-formed wings cannot fly. How these organs came into being has remained one of the mysteries of nature that needs to be enlightened."
5
"Co-optation"
Gould suggests overcoming the problem by "pre-adaptation", where a structure for one function suddenly becomes used for some totally other function, saying, "if feathers first functioned 'for' insulation and later 'for' the trapping of insect prey, a proto-wing might be built without any reference to flight."
8 However why should we expect a random insect trapper to somehow be suited for becoming an organ useful for flying? "Pre-adaptation" or "co-optation" arguments ignore the fact that functional wings are much more complex than insect trappers, as would be the case for any other such scenario: odds are against a given parent structure also being advantageous for performing some totally different unspecified daughter function which involves higher biological complexity.
However, Gould's method of explanation is widely invoked by evolutionary biologists, even at the molecular level. The National Academy of Sciences wrote:
"[S]tructures and processes that are claimed to be "irreducibly" complex typically are not on closer inspection. ... The evolution of complex systems can occur in several ways. Natural selection can bring together parts of a system for one function at one time and then, at a later time, recombine those parts with other systems of components to produce a system that has a different function. Genes can duplicated, altered, and then amplified through natural selection."14
This mechanism of "gene duplication" is a common explanation for the origin of biological complexity. The claimed advantage is that the duplicate gene gives one copy to perform the original function, and another to serve as experimental material to do something new. However gene duplications are extremely rare. The average gene only duplicates only once every 100 million years,
12 and even then, "the vast majority of
gene duplicates are silenced within a few million years, with the few survivors
subsequently experiencing strong purifying selection"
12. Another
study showed are not very free to mutate around at all, that there is strong
selection pressure on them
13. The actual mechanisms by which gene duplication contributes to evolution are not very well understood:
"However, it is unclear how duplicate genes successfully navigate
an evolutionary trajectory from an initial state of complete redundancy,
wherein one copy is likely to be expendable, to a stable situation in which
both copies are maintained by natural selection. Nor is it clear how often
these events occur."12
The bottom line is that the gene duplication explanation still leaves the
details to the dice, and this pathway definitely hasn't been experimentally
verified. This explanation is usually only based upon protein homology, it lacks a reliable mechanism, and is little better than hand waving.
Hox-Mutations or Miracle Mutations?
Some biologists have also envisioned special mutations in regulatory homeobox or "Hox" genes, where simple mutations might be able to make large developmental changes in an organism which might case a radically different phenotype. However, manipulating "Hox" genes does little to solve the problem of generating novel functional biostructures, for making large changes in phenotype are rarely beneficial. Hox gene mutations may be a more simple mechanism for generating large change, but they also do not escape the problem of the "hopeful monster":
"The drawback for scientists is that nature's shrewd economy conceals enormous complexity. Researchers are finding evidence that the Hox genes and the non-Hox homeobox genes are not independent agents but members of vast genetic networks that connect hundreds, perhaps thousands, of other genes. Change one component, and myriad others will change as well--and not necessarily for the better. Thus dreams of tinkering with nature's toolbox to bring to life what scientists call a "hopeful monster"- such as a fish with feet--are likely to remain elusive." 6
The figure below explains:
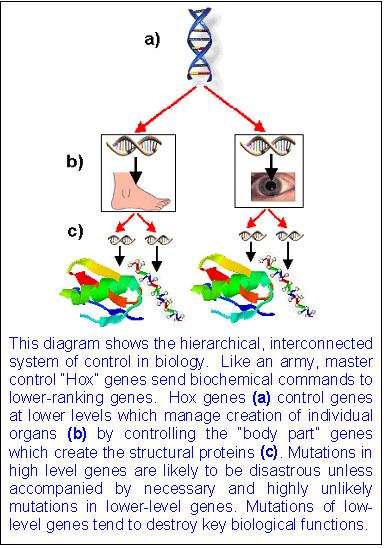
Furthermore, many biologists forget when invoking Hox gene mutations that Hox genes can only re-arrange parts which are already there--they cannot create truly novel structures. An oversimplified discussion is that genes can be thought of in two categories: "master control genes" (Hox genes) and "body part genes." "Body part genes" code for actual body parts while "master control genes" tell those "body part genes" when and where to be expressed and create their respective part. However, Hox mutations will never create new "body part genes", and thus cannot add truly new phenotypic functions into the genome, and at best we are left with the quandaries associated with "pre-adaptation". The majority of evolutionary change must take place through evolving new "body part genes", which Hox mutations cannot do. One reviewer in
Nature recognizes this fact:
"Schwartz ignores the fact that homeobox genes are selector genes. They can do nothing if the genes regulated by them are not there. It is these genes that specify in detail the adaptive structure of the organs. To be sure, turning on a homeobox gene at the wrong place can result in the appearance of an ectopic organ, but only if the genes for that organ are present in the same individual. It is totally wrong to imply that an eye could be produced by a macromutation when no eye was ever present in the lineage before. Homeotic mutations that reshuffle parts do happen, and sometimes they may have led to fixation of real evolutionary novelties, but this does not mean that such changes are implied in the majority of speciations. In fact, macromutations of this sort are probably frequently maladaptive, in contrast to the vast number of past and present species-not to mention the fact that morphological differences between related species can be minute."7
Biologist Jonathan Wells discusses the issue of Hox mutations in his book,
Icons of Evolution, where he recognizes that while Hox genes can be manipulated to cause fruit flies to sprout legs from their head. Three specific mutations are necessary to create this mutant fruit fly, and the legs are not functional, and are unbeneficial to the organism. This is a great example of why meaningful Hox mutations are complex and less simple in generating large biological change than many have promised, and how the resulting phenotype would usually be useless and disadvantageous.
Since this issue is fairly easy to understand, we'd like to just provide a couple of examples of both micro and macro-morphologies which we think are could not have functional intermediates. They defy any gradualistic Darwinian explanation, and seem to hold a level of complexity which at least very strongly implies an intelligent designer as their cause.
Biological systems for which functional intermediates seem impossible:
Major pathways of metabolism
Defense Mechanisms in Hawkmoths
The vertebrate heart
The DNA-Enzyme system
The cognitive and physiological requirements for human speech
Non-Functional Intermediates in Human Physiology
References Cited:
1. Origin of the Species by Charles Darwin
2. Lovtrup, S. [professional biologist specialising in Systematics and Developmental Biology, Dept. Animal Physiology, University of Umee, Sweden (also headed the organization of Swedish Developmental Biologists from 1979-87] (1987), Darwinism: The Refutation of a Myth, Croom Helm Ltd., Beckingham, Kent, p. 275
3. Erwin, D..H., and Valentine, J.W. "'Hopeful monsters,' transposons, and the Metazoan radiation", Proc. Natl. Acad. Sci USA 81:5482-5483, Sept 1984
4. Michael Behe, from "Molecular Machines: Experimental Support for the Design Inference" available at "http://www.arn.org/docs/behe/mb_mm92496.htm".
5. Engin Korur, "Gozlerin ve Kanatlarin Sirri"(The Mystery of the Eyes and the Wings), Bilim ve Teknik, No 203, October 1984, p. 25.
6. Nash J.M., "Where Do Toes Come From?," Time, Vol. 146, No. 5, July 31, 1995. Also at "http://www.time.com/time/magazine/archive/1995/950731/950731.science.html"
7. Book review of Sudden Origins: Fossils, Genes, and the Emergence of Species by Jeffrey H. Schwartz (Wiley: 1999). by Eors Szathmary in Nature 399:24, June 1999 pg. 745.
8. Stephen Jay Gould (1982), "Is a new and general theroy of evolution emerging?," In Maynard Smith, J. (ed.), Evolution now A century after Darwin. 129-145. Macmillan Press, London. 239 pp. First published (1980) Paleobiology, 6: 119-130.
9. Darwin's Black Box by Michael Behe, pg. 141-142.
10. Robert Carroll, Patterns and Processes of Vertebrate Evolution, Cambridge: Cambridge University Press, 1997, pp. 8-10
11. A Response to Critics of Darwin's Black Box, by Michael Behe, PCID Vol 1.1, Jan/Feb/March 2002; ISCID.org.
12. Lynch, M., Conery, J. S., "The Evolutionary Fate and Consequence of Duplicate Genes" Science 290:1151-1155 (Nov 10, 2000).
13. Huges, Austin L., "Adaptive Evolution of Genes and Genomes". (see chapter 7, "Evolution of New Protein Function" pp 143-180. (Oxford University Press, New York, 1999).
14. Science and Creationism: A View from the National Academy of Sciences (2nd Ed, 1999; NAP).







