What I did on My Summer Vacation: A Book Report and Summary of Steven Stanleys The New Evolutionary Timetable Timetable: Fossils, Genes, and the Origin of SpeciesBy Casey Luskin [Completed September, 2004; Posted January, 2005]
Preface: Steven Stanley's The New Evolutionary Timetable Timetable: Fossils, Genes, and the Origin of Species [Basic Books, 1981] (herein referred to as "TNET") is a popular-level book which develops, explains, and defends Stanleys punctuational model of evolution.[1] Stanley has been very influential in formulating the punctuational model of evolution, and he is currently a professor in the Morton K. Blaustein Department of Earth and Planetary Sciences at John Hopkins University. Stanley does not believe that the punctuational view is the only possible model for how evolutionary change takes place, but he believes it is a compelling model which yields insight into evolutionary history.
Stanley has made significant contributions to evolutionary biology, particularly with regards to elaborating the punctuational model of evolutionary change. TNET is important to understand because it provides insight into the nature of the fossil record, the punctuational model of evolutionary biology, and Stanley's own views on rates of evolutionary change and the mechanisms by which species can originate.
I should note off-the-bat that I am a skeptic of Darwinian evolution. I am an ID proponent and I am affiliated with the IDEA Center, a relatively small intelligent design organization which helps promote investigation, discussion, and debate about intelligent design theory on college and high school campuses.[2] My purpose here, however, is neither to critique nor to affirm Dr. Stanley's work, nor is my goal to make any kind of argument whatsoever, be it for or against evolution. My purpose is simply to summarize Stanleys views as presented in TNET for those interested in understanding his work. If readers disagree with what is written here, they should try to keep in mind that if I have accurately summarized Stanleys views in TNET, then they likely disagree with him, not me. My goal is essentially to attempt to provide something constructive for all sides of this never-ending debate, so that people can better understand Steven Stanley. I hope that all who read this will gain a better understanding of evolutionary biology, the punctuated equilibria model of evolutionary change, and specifically, Dr. Stanley's views.
This is loosely a chapter-by-chapter summary and review. I have skipped around a bit and combined similar sections out of the book. I have not discussed every aspect of the book, because my primary purpose is not to give every detail of Stanleys discussion, but to help readers understand his views about rates of evolution and the origin of species. Overall, I hope to accurately summarize the story told in Steven Stanley's The New Evolutionary Timetable. Introduction The first chapter of TNET contains descriptions of various terms encountered throughout the book. Evolution is an unfolding, which might occur rapidly, or gradually. Later in the book Stanley explicitly defines the gradualistic view of evolution, essentially calling it a slow and continuous, species-to-species transition which connect genera or other higher taxa. (pg. 75) Stanley immediately critiques gradualism, as traditionally embodied in the famous story of the allegedly slow, gradual evolution of the horse. (pgs. 1-3) Stanley contends that the horses evolution is actually not so gradual. Because of this, their origin is not well-documented by fossil evidence. (pg. 3) This rapid evolution is often followed by long periods of stasis, which he says is a common pattern for many of the species in the traditional story of the origin of the horse.
Stanley believes that Darwin, in The Origin of Species, diagrammed the tree of life as if branches diverged very gradually. (pg. 114) At the core of Darwins argument is the notion that species arise through gradual divergence. (pg. 115) Stanley repeatedly notes that Darwin's gradualistic views were heavily influenced by his view that the fossil record was generally too imperfect to record gradual change. Stanley criticizes Darwins metaphor of the fossil record as imperfect book, where the gradual changes were rarely recorded. This metaphor is improper because the fossil record actually says evolution proceeds by fits and starts. (pg. 7-8) Darwin believed that evolution closely follows changing environmental conditions but Stanley counters that species do not tightly track their surroundings. (pg. 11)
In Stanleys view, natural selection does occur, but it is really a combination of two processes: one random process and one nonrandom process. (pg. 11) The random component is basically comprised of the processes which create variability: reshuffling of genes in a population via random mating, plus random mutations and other genetic mechanisms.[3] The nonrandom aspect of evolution is natural selection where organisms with certain traits tend to survive better than others with different traits. One final force affecting species is genetic drift, where small populations might undergo changes which are neutral with respect to environmental adaptation but nonetheless spread throughout that population.
Though there are many definitions of the term "species," Stanley essentially would recognize a species by looking for a reproductively isolated population of interbreeding organisms. (pg. 8) Stanley firmly believes that species change little throughout most of their history. A chronospecies is a segment of a population in time which undergoes a small amount of evolution during a span of time such that the individuals during the span can be recognized as members of that same species. (pg. 14) Stanleys view of species arising quickly but then remaining generally static stands in contrast with Darwins view of the origin of species, where a population slowly morphs, via natural selection, into something else over exceedingly long periods of time.
One of Stanleys main themes in the early chapters is that Darwin taught us how changes can occur, not how species actually form. The fact that chronospecies can be identified reveals that many species change little during their history. Nonetheless, Stanley believes a chronospecies can exhibit significant variability during its overall historyprobably more than is found in a living population at any given time. Stanley also notes that one of Darwins aims in The Origin of Species was to assure the reader that he meant to use no teleology to explain how species arose. (pg. 14) Darwins scheme was totally mechanisticto an extent which surprised even Darwin! (pg. 14)
A final concept Stanley introduces is that of plenitude. Plenitude is derived from the Victorian scala naturae which held that the world was supposed to be full of life, in a gapless series filling the niches of the environment. Early evolutionists found the idea of plenitude easy to swallow because they liked the idea of species climbing some imaginary ladder of complexity. More on plenitude will come later.
A History of Darwin and his theory Chapters 2, and 3 in TNET deal mainly with the history of how Darwin developed and his theory of evolution. The story is fascinating, but it is has been told many times by many other authors.[4]
Darwin was heavily influenced by Charles Lyell, a lawyer and geologist who used arguments to ultimately convince many of his peers and antecessors that uniformitarian principles were the best way to understand geological history. (pg. 18) This view stood in contrast with the catastrophists of Lyells day.
In a dramatic encounter, Darwin experienced Lyells principles in action during an earthquake in the Chilean town of Valdivia. As the Beagle sailed north along the South American coast, Darwin saw recently-upraised beds of muscles which were putrid because they had just died after being thrust out of the tidal zone during the quake. While observing a seaside cliff, Darwin saw a pattern of muscle beds going upwards, and he realized this pattern might have been produced by many similarly small upthrusts. Darwin was intrigued by the view that the Earth is a slowly changing dynamic system and this influenced his view of lifes history as changing gradually and slowly. (pg. 19-20)
Darwin borrowed another idea from Lyellthe notion that the fossil record is imperfect. Ironically, Lyell believed life had changed little over Earths history, and cited the imperfection of the fossil record in support of this claim. Yet it was this very claim of an imperfect fossil record which led Darwin to his view of slow, gradual change.
Though the fossil record played a role in forming Darwins beliefs about extinction, Stanley notes that many of Darwins ideas about evolution were ultimately based upon studies of living organisms. Stanley insightfully divides Darwins ideas into two categories:
1) The concept of evolution (i.e. that life had simply changed over time with descent). 2) The mechanisms or processes he envisioned as causing that evolution (i.e. natural selection). (pg. 27)
Darwins voyage on the Beagle led him to think mostly about the first category: that species had changed. Stanley notes that the voyage revealed variation among organisms, and that Darwin did observe the natural struggle for survival which later influenced his ideas about natural selection. (pg. 27) But it was not until Darwin came home and read T. R. Malthus' paper on population growth that he fully began to develop his idea of natural selection. Thus, Darwin first recognized that evolution had occurred, and later came up with a way to explain how it had happened.
According Stanleys research, natural selection was born in the mind of Darwin on September 28, 1838, despite the fact that Darwin had believed in the historical concept of evolution for more than a year, though lacking an explanation. (pg. 43) Darwin soon also conceived the concept of sexual selection, which produces structures that can increase fecundity, or increase ones chances of finding a mate. The peacocks plumage or hummingbirds ornamentation are oft-cited as examples of such.
Explaining the origin of such beauty in mechanistic terms rather than as the expression of divine creativity meant that Darwin was invoking a form of argument different from those of his contemporaries. Some of Darwins arguments were explicitly dysteleological. Stanley notes that Darwin questioned why a creator would create very different species to fill similar niches in different parts of the world (pg. 27), why a single body plan would be used for species filling habitats changing greatly over time (pg. 29), or why various vertebrates would develop similarly early on and then diverge as they approached their adult form. (pg. 38) Perhaps most striking were Darwins arguments involving so-called vestigial organs, such as the pelvis of the whale or legs on some snake species. (pg. 38) Indeed, in a later chapter, Stanley also writes that Darwin complained that a Creator would never create parasites and worms which feed off the destruction of other creatures. Stanley believes this natural evil caused Christianity to shrink back from challenging the claims of biology.[5] (pg. 179)
Stanley makes two interesting observations about The Origin of Species:
Stanley here sets the stage here for his later argument as he quotes Darwin saying that natural selection requires slow, infinitesimally small modifications, but Darwin concedes that evolution might work differently if organisms could undergo more radical, sudden changes. Stanley acknowledges that Darwin did not know about modern genetics nor did Darwin consider what types of changes might be possible in small populations. But Stanley is going to put forward the view that great and sudden modifications can take place in small populations, often by mutations in regulatory genes.
Darwin thus failed to appreciate that geographical isolation of small populations might allow for rapid evolutionary change. Darwin also thought that variation was rare, and change could only occur in small, slow steps. Stanley has a different view as he believes that species are loosely packed into the environment, such that quite a bit of variation can be tolerated. Because there is this latent variation, small and isolated populations might trap some of it, permitting rapid emplacement of new traits and divergence into new species. (pg. 53) In Stanleys view, natural election does operate, but in a manner that is much less constrained than how Darwin imagined. (pg. 53)
Stanley speculates that it took Darwin 21 years to publish Origin because Darwin first wanted to establish a reputation as a scientist before putting forth challenging ideas. In 1849, Darwin wrote that he was only writing in a speculatory manner. (pg. 59) Yet Darwin was very good at swaying skeptics, of which he had many, partly because he was very sensitive to criticism. When criticized, Darwin would emphasize where he and his critics shared common ground, rather than where they differed.[6]
Darwin sought specifically to convince three prominent British scientists: Charles Lyell, Joseph Hooker, and Thomas Henry Huxley. Though Lyell was always skeptical, he encouraged Darwin to publish. Hooker eventually accepted evolution (pg. 57), and Huxley became a staunch supporter. Yet Stanley believes Huxley questioned whether or not gradualism was correct and thought there may have been more rapid steps in evolution. Despite these differences, when Darwin retreated from public debate after publication of Origin, Huxley became one of his most ardent proponents.
Stanley states that the strongest opposition to Darwin was religious, claiming that even Lyell may have objected to Darwin for religious reasons. This was probably because, in the minds of many, Darwins work demoted the status of humans from Gods special creation to the level of lower mammals. (pg. 60) Stanley summarizes the reaction to Origin saying that most natural scientists contemporary with Darwin were impressed, and many accepted the historical concept of evolution while remaining skeptical that natural selection was its mechanism. (pg. 60) Darwin's Theory after Darwin Chapter 4 of TNET discusses the history of Darwin's theory subsequent to Darwin. Evolutionists after Darwin exerted much energy trying to elucidate the shape of the tree of life, because they accepted that evolution had generally taken place. But in the 1880s, Darwins ideas were again questioned when Kelvin argued that the Earth was so hot that it could not have been more than about 75 million years old--an age of the Earth which was younger than what many had expected as necessary to accommodate Darwins account of evolutionary history.[7] Although essentially nothing was known about population genetics and mutation rates at this time, Kelvins ideas had the effect of casting doubt upon gradualistic evolution. It was not until the discovery of radioactive heating in the early 1900s that a source for the Earths heat was identified, allowing the mainstream view of the age of the Earth to increase, and apparently increasing the confidence of biologists in gradualism.
Another obstacle for Darwins mechanism of natural selection was explaining the origin of variation. Darwin's contemporaries believed in a vague notion called blending inheritance where the traits of parents would somehow be blended in a non-discrete manner into their offspring. Under this model, any new adaptation would eventually get diluted out by being mixed during inheritance with other structures. (pg. 62) This was a severe source of doubt towards Darwins theory early on, even for Darwin himself. It was not until the turn of the 20th century that geneticists began to accept Mendelian genetics, where traits were inherited as discrete entities (i.e. alleles) such that an allele could persist for generations. Only then did biologists find a renewal of confidence that natural selection could allow for permanent biological changes in species.
Darwins mechanisms for gradual evolution by natural selection were again questioned in the early 1900's with the advent of the macromutation. (pg. 66) If mutations could cause great phenotypic changes, then gradual selection wasnt so important for there was no longer any need for gradual accumulation of small beneficial mutations. (pg. 66) Macromutations were viewed as a panacea for evolution. Stanley notes that macromutationists caused quite a stir and many of this period even adopted Lamarckianism.
In the 1920s and 30s, the Modern Synthesis of evolutionary biology was born, and biologists came back to a gradualistic view of evolution by natural selection. (pg. 67) Macromutations fell out of favor as geneticists learned that small changes in hereditary molecules (i.e. what we now know to be DNA) indeed really could cause miniscule phenotypic changes. This made it possible for evolution to be explained by the occasional small and beneficial mutation, leaving less of a need to invoke macromutations.
In the 1950s, Theodosius Dobzhansky observed small changes in fruit flies, producing traits which he could then select for in the laboratory. This meant evolution could be studied in the laboratory, elevating it from a historical and conjectural field to a more rigorous experimental one. (pg. 68) The growth of evolutionary biology as a scientific field was also aided by the development of the mathematics of population genetics.
Sewall Wright made key mathematical findings which aided population genetics. Stanley believes Wrights work supports the important concept that small populations are more widely subject to chance changes in gene frequencies while large populations are not good at fixing such changes. (pg. 68, 69) Essentially, Wrights findings explain the common intuition that when dealing with smaller numbers, things become more unpredictable. (pg. 68-69) When a mutation arises in a smaller population that is isolated from the larger population, it has a better chance of becoming fixed into more individuals. Dobzhansky applied these ideas under the context of Darwins gradual changes, where a sub-population with a favorable mutation slowly spreads its mutation into the larger population over long periods of time. Yet the possibility of rapid speciation was not considered by Dobzhansky.
Stanley believes that evolution will proceed more slowly in large populations because it is difficult to fix a new trait over a large geographical range. (pg. 70) Rapid evolution is most likely in small isolated populations which have well-mixed breeding, perhaps even inbreeding. These populations will likely occupy a restricted habitat where there might be novel and strong selection pressure. (pg. 70) This can permit unique scenarios for evolutionary change. Stanley notes that there was little room for dissent under the guards of the Modern Synthesis as many who proposed rapid evolution were ignored or ostracized.
Ernst Mayr had envisioned something like a punctuationist model in 1942 when he proposed that some living genera of birds evolved rapidly out of small, peripheral populations diverging from the original population. (pg. 77) Mayr believed this allowed him to better explain the sudden appearance of new morphology. (pg. 77) Of important note is Stanleys observation that if a transition is rapid and the population small and localized, we would not expect to find fossil evidence of the transformational event. (pg. 77) Stanley thus believes this model best accounts for the sudden appearance of many species in the fossil record. This also implies that the lack of gradual transitions is not simply the result of an imperfect record, because most species typically change very little throughout their history. Mayrs ideas were apparently generally ignored by geneticists who saw evolution progressing extremely gradually. (pg. 78)
In the 1970s, Eldredge and Gould revived Mayrs ideas. Stanley believes that polarization of viewpoints and misunderstandings have marked the field since that time. Gradualists misunderstood the punctuational view in that they thought it denied that gradual evolution ever occurs within established species. Similarly, Stanley believes punctuationists like Gould and Eldredge have wrongly assumed that the gradualistic model taught that change was never rapidly divergent. Stanley proposes that there should be two different models, the gradualistic model and the punctuational model, which need not be mutually exclusive. According to Stanley, both models talk about different, but real, modes of evolution.
Exploring the Fossil Record In chapter 5 of TNET, Stanley turns to the fossil record to explain lifes history because it can provide direct evidence of large-scale evolutionary change. (pg. 72) The fossil record can allows us to test gradualism, but Stanley believes the verdict is against gradualism, and he give numerous examples to support this contention.
According to Stanley, there are numerous examples of species which remain unchanged for long periods of time. The best tests of the two models come from recent fossils because here we have more complete fossil records to work with, including some collections leading all the way up to living representatives. Such lineages are claimed to vindicate stasis as the common mode of existence for species. Examples of living fossils which exhibit stasis include: - Ice Age beetles being almost identical to modern ones (pg. 79) - Bowfin fish since the Cretaceous (pg. 83) - Lungfish (pg. 84) - Sturgeon fishes (pg. 84) - Garpikes (pg. 84) - Snapping turtles (pg. 84) - Alligators (pg. 84) - Tapirs (pg. 84) - Aardvarks (pg. 84) A similar pattern is found in extinct mammals from the Bighorn Basin in Wyoming, which evolved very little during their duration. (pg. 85-86) Recent mammals such as the wooly mammoth and wooly rhino show a similar pattern, in that after arising, they evolved little. (pg. 98) Another conundrum for gradualism comes from Stanleys observation that the Pleistocene period shows that during a relatively short period, at least fifteen new genera of mammals appeared in Europe.
Stanley characterizes the origin of mammals as an adaptive radiation, which is basically the rapid divergence of many species from a common ancestor. (pg. 86) During adaptive radiations, species quickly explore available niches. (pg. 95) The advantage of invoking adaptive radiation is that biologists are no longer required to conjecture unrecorded periods of evolution or claim that the fossil record is simply incomplete. (pg. 89, 90)
The mammalian radiation at the beginning of the Cenozoic was made possible by the sudden extinction of the dinosaurs which opened up various niches. Stanley notes Mesozoic mammals were small and rodentlike (no bigger than a house cat) yet in about a twelve million year period, most of the living orders of mammals had come into existence. (pg. 91, 92) Among the orders which appeared rapidly were those which include bears lions, wolves, bears, horses, rhinos, deer, pigs, antelopes, sheep, whales, and bats. Stanley sees one of the greatest challenges posed to gradualism coming from the origin of bats and whales, which he says evolved from terrestrial mammals in a time-period less than twelve million years. (pg. 93, 96)
In Stanley's view, the greatest adaptive radiation was the Cambrian explosion. (pg. 86) Darwin knew about the relatively sudden appearance of the Cambrian fauna but proposed that the record testifying to their gradual evolution came lie in sediments which were inaccessible. (pg. 87) Stanley believes that the Cambrian explosion is a real phenomenon, but also discusses various trace fossils which have been discovered in late Pre-Cambrian sediments. Nonetheless, the Pre-Cambrian life lies only in the strata just below the Cambrian, implying there was rapid evolution. (pg. 89) Stanley claims that gradualism cannot explain the relatively rapid origin of these groups, which thus requires punctuational evolution. (pg. 90)
Stanley notes that similar evidence comes from the origin of angiosperms, where many species also appear suddenly. Like the Cambrian fauna, Darwin also knew about the sudden appearance of angiosperms and was perplexed by it. (pg. 90) Stanley believes that the fossil record shows a progressional increase in complexity of plants with respect to leaves and pollen, but concludes that this evolution was fairly rapidit took place in about ten million years. (pg. 91) Stanley believes this too challenges Darwins gradualism.
To test the gradualistic vs. punctuational models, Stanley asks if there is enough time for the gradualistic model to account for the origin of bats or whales. According to the fossil record, a small rodentlike mammal transformed into a bat, and also to a whale in about ten to twelve million years. If we dont invoke rapid branching, then we must rely upon gradual transformation. However, chronospecies of Cenozoic mammals typically exist for more than a million years before they exhibit enough change to be considered a new species. This only gives about ten to fifteen chronospecies with which to go from a rodent to a bat, and a rodent to a whale. Because differences between successive chronospecies typically only slight, there is simply not enough time in the fossil record to allow for this change under the gradualistic model. (pg. 93-94) Stanley thus believes this implies that the punctuational model must be invoked to account for the origin of bats and whales.
To account for the fossil data, the gradualist is forced to assert very rapid evolution within established species. This means that the chronospecies must have been of very short duration. But, just as in the Cambrian and angiosperm situations, there is no evidence that chronospecies have undergone rapid change. (pg. 94) A problem faced by the gradualist is thus to explain why an established species would undergo rapid change, and why the mere presence of open ecological niches would cause an entire established species to abandon its habitat for a new mode of existence? (pg. 94) We would not expect an entire desertion of what worked just fine before. (pg. 94) Stanley proposes that under the punctuational model, this rapid diversification occurs by the sprouting off of new species from already established species, where the original species can continue on in its old mode of existence. (pg. 94) The availability of ecological niches can drive this rapid branching process, and this is what has happened during adaptive radiations. Stanley refers to this solution as "quantum speciation." (pg. 96) Thus, Stanley proposes a solution to the origin of whales by postulating that there was never a single species which gradually morphed from one form into the next until it eventually turned from a rodent into a whale. Rather, very small segments of the chronospecies would branch off and "sprout" into something new, while the rest of the original population of the chronospecies would continue to do its original thing (figure 1):
Similarly, to solve the problem of the origin of so many new genera during the Pleistocene, Stanley notes that there is only enough time for three or four successive chronospecies, which might produce a new genus. Under the punctuational model, Stanley believes new genera might arise after one or two speciation events. (pg. 99) Stanley speculates that some genera, such as polar bears, are sufficiently similar to other species (i.e. the brown bear) that they might have formed in just one speciation event. Thus, we have no choice but to invoke the rapid speciation of populations which were too small to leave a record of their evolution. (pg. 99)
The record of elephants provides another example of a punctuational pattern. Each modern elephant genus appears rapidly from its ancestral genus Primelphas, and then undergoes little change for thousands of generations. (pg. 100) This clearly shows the punctuational pattern.
Another confirmation of the punctuationist model comes from formanifera, which are tiny one-celled marine creatures that live in seafloor sediments. Stanley notes that the average Cenozoic bottom-dwelling foram species exists for about twenty million years and undergoes very little change. (pg. 100) The fact that thirty families of forams appeared in the first 30 million years of the Cenozoic is significant because it points against the possibility that they arose due to the transformation of already-existing species. (pg. 101) Bivalve mollusks also exhibit a punctuational pattern for their origin.
Stanley notes that he is by no means the first paleontologist to uncover evidence that conflicts with gradualism. Some paleontologists from Darwins era uncovered paleontological data conflicting with gradualism, but they were generally ignored. T.H. Huxley believed the origin of every class must have been much older than its first appearance in the fossil record and postulated hypothetical lands which hosted long, but undocumented periods of evolution. (pg. 103) Darwin again assuaged these doubts by appealing to the incompleteness of the fossil record, but Stanley writes that eventually scientists forgot that Darwins views about the incompleteness of the fossil record were not based on solid evidence of widespread gaps, but merely upon deduction from his gradualistic model. (pg. 106) This unfortunately led to circular reasoning with regards to the incompleteness of the fossil record (pg. 106)
Stanleys makes an argument that if we see a pattern which doesnt fit one model, then we must abandon it in favor of another model. The data shows that gradualism isnt what happened, so we must accept that most change occurred in small populations permitting the punctuational pattern. (pg. 109) Darwin could not have proposed this because it would have meant the evolution he proposed occurred exactly where we would be least likely to observe it. Such an idea, even if correct, would have been difficult to test and thus unlikely to gain the favor of many scientists. (pg. 110)
Rapid Species Formation In 1970, Ernst Mayr predicted that evolution would one day rely heavily upon the changes in regulatory genes. (pg. 108) Stanley believes that changes in regulatory genes, which guide the development of an organism, are key to the punctuational model.
Today, Stanley believes the punctuational model is more reasonable because today we have living examples of species that have formed rapidly. In fact, Stanley states that contrary to what Darwin believed, only a small percentage of genera formed via gradualistic transformation of an entire species. (pg. 114) Gradual evolution might still occasionally happen if two conditions are met: 1) Natural selection is consistent throughout the species range for long periods of time. (pg. 132) 2) Similar ranges of variability are widespread throughout the population. For some types of changes, like changes in body size, this sort of gradual evolution may be possible. According to Copes Rule, species tend to get bigger with time, for there seems to be a more-universal selection pressure allowing larger animals to better find food, to fight off predators, and to outcompete others vying to mate. (pg. 133) Occasionally, Stanley speculates that changes related to reproduction might also find more ubiquitous selection pressure. This could be able to account for large-scale sexual selection for certain visually attractive traits, such as the male peacocks tail. (pg. 133) However, for many other traits, this second criterion seems difficult to meet, particularly for unusual genetic changes, which might play a large role in the origin of species. (pg. 132)
To place confidence in the punctuational model of evolution, Stanley seeks examples of speciation which meet two criteria: 1) It must have occurred rapidly (i.e. in less than a few thousand years or at most a few tens of thousands of years). (pg. 111) 2) It must have achieved significant morphological change (such as change leading to the formation of a new genus). (pg. 111) Quantum speciation or quantum evolution represent the notion that there has been rapid evolution in a small population. (pg. 115) Quantum speciation had previously been given little attention because it was associated with Goldschmidts hopeful monsters, judged to be an untenable hypothesis by biologists. (pg. 135) Quantum speciation events take place in special environments, which may be difficult to identify because they are short-lived or become obscured if the species becomes widely dispersed from the local environment where it originated. Thus, a third necessity for finding such an example of quantum speciation may be that the species has not yet escaped from their place of origin. (pg. 111)
The first example Stanley gives is that of African cichlid fishes in Lake Nabugabo in Uganda. This small lake was separated from nearby Lake Victoria within the past four-thousand years, and contains five species of cichlids which are unique to the lake. Each species is similar to its parent species in Lake Victoria, typically differing in coloration and other minor traits. (pg. 111) (Changes in male coloration can be key to establishing reproductive isolation because this is how cichlids typically identify members of their own species for breeding.) Thus, here we find an example which meets criteria 1rapid speciationin that reproductive isolation may have been achieved quickly. It may not meet criteria 2, but it does show that a reproductively isolated population a species can be formed rapidly. (pg. 111)
Stanleys second example comes from Hawaiian moths of the genus Hedylepta. Species in this genus feed on many plantsranging from grasses, to lilies, palms, legumes, and banana plants. Yet bananas did not arrive in Hawaii until about 1000 years ago when brought by the Polynesians. Stanley believes multiple speciation events must have occurred leading to reproductively isolated populations of moths which feed exclusively on the banana plants in the mountain forests. Again, effective reproductive isolation has been established with these moths, and thus they are a species. A well-known example Stanley discusses is the peppered moth, which is said to have undergone a rapid coloration change. However, this was not significant enough morphological change to constitute even a speciation event. (pg. 130) These examples seem to meet criteria one (rapid change). But what about criteria twosignificant morphological change?
Stanley finds evidence for rapid significant morphological change from analysis of fishes and birds. In the Death Valley region of California, desert pupfishes populate recently produced ponds which are isolated from other ponds. Some of these recently produced ponds contain unique species of pupfishes, one of which has reduced pelvic fins and does not resemble other pupfishes. Honeycreeper birds on the Hawaiian Islands arose from an ancestral population of finches and today display widely diverse beak shapes. (pg. 115-116) There are honeycreepers with parrot-like beaks, long curved beaks, nondescript beaks, and feed on a variety of different food sources. Some of these species are endemic to the big island of Hawaii, identified as only 750,000 years old. Stanley believes these could be examples of the rapid origin of significant morphological change.
A final example comes from Lake Victoria in Africa which contains cichlid species with many different types of feeding apparati. From these cichlids, Stanley finds an ideal test between the gradualist and punctuational models. The many cichlid species in Lake Victoria are said to have descended from a riverine species which entered the lake sometime after it formed. The gradualist model would predict that the riverine species, once in the lake, would have slowly morphed into another species, such that examples the original riverine species ceased to exist because they were slowly selected out over time. Yet the riverine species persists in Lake Victoria.
Furthermore, Stanley notes that the average fresh-water fish chronospecies takes several million years to evolve into a new chronospecies. Yet there are 170 unique species of cichlids in the relatively young lake. Gradualism cannot account for this high species diversity. But under the punctuationist scheme, this is easily explained because the segments of the riverine species population which entered the lake rapidly budded off into other species. (pg. 119; see also figure 1.) These cichlid species illustrate how adaptive radiations can take place when there is ecological opportunity and environmental niches can be filled.[8] Ecological opportunity is thus important to quantum speciation.
Adaptive radiations typically take place in environments with clear boundaries (like lakes or islands), such that their inhabitants are geographically isolated from other species. Yet geographical isolation does not care about the scale of spatial separation. (pg. 122) All that matters is that there is some physical separation. Speciation events often include passage through a population bottleneck. Stanley believes that population bottlenecks do not represent the transformation of an entire species, but rather represent a small localized segment of a larger population which is rapidly transformed.
But what about the origin of all the variation which permits rapid speciation? Stanley attempts to account for this by recounting the story of the artificial introduction of Bairdiella (a small fish) into the Salton Sea in the early 1950s. Bairdiella was the only fish species in this lake when it was introduced. After its introduction, members of subsequent generations often had abnormalities and deformities. But once the population filled the lake and food resources became scarce, the odd diversity disappeared. This illustrates stabilizing selection where competition tends to weed out abnormalities. (pg. 121) However, the presence of that diversity in the absence of competition illustrates that much variation might be available when there is less competition. This, Stanley believes, reveals that much unseen potential variability exists even during normal environmental conditions. (pg. 121) Some habitats with relaxed selection may not weed out that variability such that new changes can occur. (pg. 122)
Stanley also looks to the fossil record to support these views. When the Himalayas were formed about 10 million years ago, an adaptive radiation of hydrobioid snails took place due to the creation of a large habitat. A similar scenario occurred when marsupials invaded Australia. Again, the availability of ecological opportunity yielded an adaptive radiation among marsupials. Stanley calls these geographically isolated events insular adaptive radiations. (pg. 126) Yet because they often occur in permanently isolated environments, many such insular radiations go unrecorded because their products eventually go extinct, leaving no descendants. Yet Stanley believes that quantum speciation is going on constantly, sometimes as insular radiation and sometimes simply as non-isolated local events. (pg. 126)
To provide an account for the origin of variation in the first place, Stanley contends that small genetic changes can indeed cause great morphological changes. Stanley attributes the origin of this variation to changes in regulatory genes, which are those which switch on-and-off structural genes. Structural genes are those that form the proteins which form physical structures in the cell, and which mediate biochemical reactions. (pg. 127) By manipulating structural genes, Stanley believes small changes in regulatory genes might have large effects on morphology.
Chromosomal mutations or rearrangements might also play a role in quantum evolution, especially if the operation of a regulatory gene is affected by its position on a chromosome relative to other genes it could potentially control. (pg. 131) The fact that many closely related species or genera differ in chromosomal structure, indicates that chromosomal rearrangements could cause evolution. This must take place in small populations because parents with different chromosomal arrangements usually do not produce successful offspring. (pg. 131) For new chromosomal arrangements to spread, an organism must be in close proximity to other individuals with the same chromosomal structure. Inbreeding in a small population would best facilitate such a scenario.
These types of mutations might change the sequence of development or the rate at which structures grow. (pg. 131) Stanleys primary example given of the power of changes in regulatory genes comes from the axolotl, and amphibian which has arrested development due to a mutation which prevents production of hormones controlling development. Yet injection of the missing hormones causes the amphibians to mature fully and become land-dwelling adults. Stanley believes that a single mutation in to this developmentally important hormone could have caused the origin of this species.[9]
A second example comes from the giant panda, which has enlarged and strengthened teeth and jaws, but a reduced size in genitalia and the pelvic area. Stanley claims that simple changes in growth gradients in a bear could result in these characteristics. Given the sparse fossil record of giant pandas, Stanley imagines that they may have arisen in a local bamboo forest high in the Himalayas in a relatively short period of geological time.
A final hypothetical example discussed by Stanley where regulatory genes might have played a large role in evolution comes from snails. Stanley first notes that a common objection to the idea that rapid evolutionary change is possible in small populations is that it facially appears unlikely that an individual with many new traits could find a mate. (pg. 133) Stanley claims first-off that this objection comes primarily from biologists (and not paleontologists) who misunderstand the vastness of geological time. He believes that snails can help overcome this objection.
During their development, snail bodies undergo torsion such that their bodies are twisted so the anus lies above the head. (pg. 133) This torsion occurs by the differential development of two muscles in juvenile snails. In some species the process takes only a few minutes (pg. 133) Stanley claims that the new morphology caused by torsion could evolve rapidly. He finds evidence for this from the fact that when snails have lost torsion, they have often not lost it altogether, but to varying degrees. Thus detorsion has evolved by in progressive steps. But because torsion itself is not found developed in varying degrees, he believes it must have arisen in a single mutational step. (pg. 134)
Stanley seeks to find out if this sort of mutation would have been possible in the ten million years preceding the Cambrian Explosion. In the Pre-Cambrian pre-snail (i.e. snail-like organism with no torsion) population, it is conceivable that each year there could be as many as a hundred-thousand-trillion broods of presnails. With such numbers, it is easy to imagine that some snails would have the single mutation required for torsion, if such a single mutation exists. But what about the odds that two members with the correct mutation would find one-another? Stanley says that this is possible through inbreeding. If a female had a mutation in her gonad tissue such that many of her offspring had this particular mutation, then again, chances are high that some of her own offspring would find one-another and reproduce, producing more offspring with the single torsion mutation. If some of these siblings happened to experience geographic isolation from the rest of the population, reproductive isolation could be established. (pg. 134) Thus, Stanley concludes that torsion could arise rapidly, because with a high-fecundity type of organisms like snails, the odds are good of getting a single mutation fixed in less than ten-million years.
One implication of Stanleys argument is that fixation of new traits might require inbreeding. This is especially true when the new trait is recessive and members in a large species carrying genes for that trait would be unlikely to ever meet and breed. A small local population would thus be necessary to allow for the trait to be readily expressed and allow natural selection to take effect.
One difficulty Stanley identifies is pinpointing the exact moment in when a speciation event takes place. Stanley suggests that the definitive moment of speciation is when a population can no longer interbreed with its parent population. This moment, however, is difficult to identify. Reproductive isolation might not always make a good marker, for the rapid evolutionary changes which typically mark speciation may occur in a population which is already reproductively isolated, or one which is not completely reproductively isolated. Stanley isnt worried about this too much, for it is difficult to identify the exact moment of speciation, even if it ultimately takes place. (pg. 132) Finally, it is difficult to give general answers about when exactly speciation occurs because modes of reproductive isolation may differ from species to species, depending upon the lifestyle, habitat, behavior, and mobility of a type of organism.
One final important note is that many of the examples Stanley gives for rapid speciation represent small populations which could easily be environmentally threatened. Such populations are of great value to the study of biology, however because of their small size, they are often not widely recognized as important examples of biodiversity. Environmental protection is vital for these sensitive species which could hold keys to understanding the punctuational mode of evolution. Stanley doesnt view all organisms as connected in a web, and he isnt sure that removal of one species will always affect other species. But he believes environmental protection is still necessary because of aesthetic sensibilities, the sanctity of life, and potential scientific or educational value. (pg. 197)
The Punctuationism and Humanity Stanley discusses human origins because he believes that although gradualism has dominated this field, a punctuational pattern best explains human origins.[10] Traditionally, anthropologists and paleontologists have reconstructed human evolution as an ape gradually becoming human. (pg. 138) This gradualistic scheme has dominated thinking in evolutionary biology since it was first promoted by Darwin and Huxley, where one species gradually morphed from an ape species into humans. This is referred to as the single-species hypothesis where humans become the greatest achievement of evolution. (pg. 164) Stanley seeks to have that view overturned with regards to human origins.
One way to test between the punctuational and gradualistic models is to look at the branching patterns. A branching pattern is required under the punctuational model, but it is does not constitute proof of the model. The decisive question is if the hominid species leading to humans evolved via quantum speciation with rapid divergence, or gradual transformation. Stanley finds validation for the punctuational model from the fact that many hominid species are long-lived. He believes our genus evolved from asutralopithecines in just a few million years, but yet most hominid species live up to one-million years. Stanley provides a summary of various hominid species which have existed in the past 4 million years:[11]
Australopithecus africanus is the species Stanley believes was the direct ancestor of the genus Homo. This slender australopithecine species lived at least from 3 to 2 million years ago (Ma), and perhaps as late as 1.6 Ma. Humans have an average brain size of about 1300 cubic centimeters (cc) while Australopithecus africanus has an average brain size of about 500 cc. Stanley does note that brain size of an individual is not necessarily a reliable indicator of intelligence, because brain size within a species can vary greatly without corresponding to intelligence. Nonetheless, the differences between humans and australopithecines is great enough to show a significant intellectual change. (pg. 145) Other differences between humans and australopithecines include height (humans are about a foot taller), forehead size (ours is taller and more vertical), and the fact that we have a chin, reduced orbits, jaws, and teeth, and we have upper limbs with higher dexterity.
Australopithecus afarensis is also a slender Australopithecine, but older than A. africanus. A. afaraensis left footprints about 3 Ma that seem to imply a mode of locomotion similar to modern humans. Stanley thinks it is very similar to A. africanus such that they could be united as a single species. A third type of australopithecine Stanley mentions are the robust form, but he does not believe their line led to humans. (pg. 151)
Stanley recounts that Homo erectus appears about 1.6 Ma and lived until at least 200,000 year ago. Stanley believes erectus is closely related to Homo sapiens.[12] Stanley notes that Homo erectus has jaws, teeth, and braincase which are of a size between that of humans and the australopithecines. Brain volume in Homo erectus ranged from 800 cc to 1300 cc, with the upper limit within the range of average variation in our own species. However, because they lacked the high forehead it can be inferred that modern humans are more intelligent. (pg. 146) H. erectus had an upright posture differing little from modern humans and probably used fire and various stone tools.
When introducing Homo habilis, Stanley notes is controversial because some paleontologists do not believe it exists. (pg. 148) Its specimens are dated between 1.8 and 1.6 Ma. Stanley questions if habilis should be in the genus Homo because in many aspects of skull shape and size, habilis seems like the australopithecines. (pg. 148) It seems to also have used crude stone tools.
Neanderthals, Stanley believes, had unique features not found in earlier populations nor in modern humans. This makes it difficult to determine whether or not they were our ancestors. Neanderthals lasted at most 100,000 years and had brains slightly larger than those of modern humans. They also had a stockier build than modern humans with heavier limbs, and were thus probably physically stronger. Stanley also notes some relatively small differences in skull shape, chin shape, and pelvis shape. Culturally, they made knives and other sophisticated stone tools. As for their classification, many researchers have called them a member of our own species but Stanley believes these interpretations may have been colored by biases from gradualism or a desire not to have another species in Homo with a larger brain size than modern humans.
As for our own species, Homo sapiens, Stanley believes we are even more specialized than Neanderthals with regards to our sharp chin, weak brow, and highly vaulted forehead. We also have culture more highly developed than other members of Homo.
Stanley uses habilis as a springboard for talking about how the origin of the genus Homo works best under the punctuationist model. Because some australopithecines may have existed as late as 1.6 2.0 Ma and Homo erectus definitely lived by 1.6 Ma, Stanley infers that there is simply not enough time allowed in the fossil record for a gradualist model to accommodate Australopithecus africanus turning into Homo habilis and then turning into Homo erectus.
But how did Homo sapiens arise? Gradualists who believe in the single species hypothesis believe that Neanderthals were members of Homo sapiens. As noted, Stanley believes this classification may be improper, and has been the result of a fear that a hominid species other than our own might claim the title of the largest brain. Stanley also believes that skeletal differences between Neanderthals and Homo sapiens are not insignificant, and that in other taxa of mammals, such differences would warrant the creation of a new species. Finally, our cultures appear to be quite different. Stanley states the Neanderthal culture did not go much beyond making particular types of stone tools. But the advanced artistic culture of Homo sapiens rapidly arose all around Europe between about forty thousand and thirty-five thousand years ago. Thus, he believes Neanderthals should be called Homo neanderthalensis, a species distinct from our own.
The punctuational model should be invoked to explain the origin of Neanderthals and Homo sapiens. According to Stanley, Neanderthals appear about 100,000 years ago and disappear perhaps just a bit later than modern humans appear. Stanley says that Neanderthals could not have transformed into modern humans so quickly. (pg. 153) Neanderthals had existed for at least 65,000 years with basically no change, and then suddenly Homo sapiens appears. Stanley claims this could only have happened if a small population of Neanderthals had undergone a quantum speciation event. (pg. 153)
Apart from paleontological timescales, Stanley also attempts to account for the origin of many of our traits in a punctuationist manner. He believes many human traits can be explained through the retardation of development, called neoteny. (pg. 156) Firstly, Stanley believes that many of our modern human features resemble those of newborn apes: our generally hairless bodies, our soft hands, and our enlarged, high-vaulted braincases. Stanley believes the result of these traits is that our newborns are more helpless and must be cared for and trained, causing more social interaction, ultimately resulting in a more elaborate culture. Stanley believes that a neotenous pattern of development could also explain the origin of upright posture. Stanley prefers this mode of change because he believes that neotenous changes in the developmental program might take place with only a few mutations, making rapid morphological change more likely.
Interpretation of phylogeny can be very different under the gradualist vs. punctuationist model. Stanley notes that due to the gradualist model, fossils are placed in a phylogeny based upon the assumption that each population is a bit closer to becoming a human. (pg. 151) Because of the rapid morphological changes permitted under the punctuational model, such assumptions can be thrown out. Speciation does not mandate that a new species follow an evolutionary trajectory of previous species because during quantum speciation, changes may be the function of accidental selection pressures in the local environment or random mutational events. (pg. 151) There is thus no progression towards one form (like Homo sapiens). (pg. 151)
Punctuational evolution is not phased by evolutionary reversals such as the fact Homo sapiens has a brow which is less robust than that of Homo erectus, despite the fact that Homo erectus is more heavily browed than the australopithecines. (pg. 151) Because of the ease with which species can change the direction of evolution under the punctuational model, Stanley argues that the fact that Australopithecus africanus is specialized does not disqualify it from being a direct ancestor of habilis or the genus Homo. Thus, he would argue that the punctuationist model can easily account for the differences between africanus and Homo. The fact that the two slender australopithecine[13] species change very little over hundreds of thousands of years makes it improbable that a gradualist model can account for the origin of Homo.
Looking at this pattern from a broader perspective, Stanley believes that hominids have not really evolved any faster than many other mammalian taxa. It is thus a false interpretation to search for the missing link because the notion that two species must be connected by species of intermediate form is not a requirement under the punctuational model. (pg. 155) This is validated by the presence of evolutionary reversals. Evolution does not necessarily move only in one direction. (pg. 155)
As far as our own cultural evolution goes, Stanley believes that the culture of modern humans appeared very rapidly in Europe about 35-40 thousand years ago. Modern theories of social evolution, which focus more on the interaction of groups might do better to draw upon ideas from punctuational models of evolution rather than gradualistic models. (pg. 205) One analogy can be drawn from the fact that species normally tend to resist change, a trait often found in large governments or corporations. Yet new kinds of social structures may arise under small localized conditions, such as the United States during the American Revolutionary period. (pg. 205)
Finally, Stanley believes that humans are not evolving much anymore. Medical advances have allowed many who in the past were not allowed to breed to do so. But since modern humans arose in Western Europe, he says there has been essentially no skeletal evolution. Furthermore, he observes that humanity tends to reject mutants whether natural mutants or potentially engineered ones and this could retard future evolution. Stanley believes that if major adaptive genetic changes occur during speciation, then speciation probably wont occur within modern humans unless we escape Earth and colonize other planets. Or, more glibly, if nuclear war were to decimate much of humanity, then the subsequent populations might also cause a new race to come about.
More Social Commentary In some of the final chapters, Stanley discusses his take on creationism. By creationists, Stanley seems to mean almost entirely young Earth creationists who argue their position based primarily off of the Bible.[14] He indicates he would prefer that creationism just be ignored, but because of its influence upon education, it cannot be. (pg. 177) Yet, Stanley takes a broad view of divine creation arguing that it represents, on one end of the spectrum, a 6-day Genesis account (where the rock record is the result of Noahs flood), and on the other, progressive creation where species are created over long periods of time. Either way, Stanley sees these views as incompatible with science because they are both subject to the will of God, which cannot be studied by scientific methods.
Stanley observes that evolutionists represent a wide range of religious viewpoints, whereas creationists are primarily composed of Christians. Some evolutionists are Christians while others practice other religions, or they are agnostics or atheists. Stanley writes that T.H. Huxley coined the term agnostic which means having no knowledge with regards to the existence of God. (pg. 167) As far as science is concerned, Stanley writes that agnosticism is the only acceptable position regarding the existence of God. According to Stanley, atheism is just as inappropriate as a scientific conclusion as is theism. (pg. 167) Stanley then juxtaposes scientific knowledge and religious knowledge where observations and measurement are the stuff of scientific knowledge and revelation and faith are the basis for religious knowledge. Because of these differences in knowledge, Stanley believes that religion and science occupy separate domains. (pg. 167)
Another difference between religion and science is the standard of proof. In science, things never reach absolute proof. One only achieves degrees of support through the data. Stanley characterizes creationist opposition to evolution as if it merely asks the question, has evolution been proven? Stanley answers that most evolutionists would argue that it evolution is a real phenomenon, but absolute proof in science is impossible. (pg. 169) Stanley believes that most nonfundamentalist biologists are evolutionists, because evolution has not been disproved. Stanley finds 2 ways a theory can be disproved: 1) Discovery of direct evidence opposing the theory 2) Refutation of its corollaries or predictions (pg. 171) With regards to evolution, Stanley believes that evolution makes many predictions, but no prediction of evolution has been refuted to the degree that it has lost general support. (pg. 171) Stanley then lists some of the ways evolution might have been disproved: - Discovery of a fossil population of horses in Pre-Cambrian rocks - Any topsy-turvy sequence of fossils - A biogeographical inconsistency (Stanley proposes as kangaroos found in California identical to those in Australia) Stanley believes that evolution has also solved new problems, such as the fact that some odd geographical distributions were resolved in light of plate tectonics. Stanley claims that essentially the existence of any of transitional forms would refute a creation account. He offers Archaeopteryx and ammonoids as such examples, claiming the evolution of nautiloids into ammonoids is recorded in the fossil record, particularly with regards to the position of the tube in their shell.
Stanley believes that his views will make some creationists happy because the punctuational model teaches that some species appear suddenly in the fossil record, and then change slowly. Yet Stanley turns this observation into fortune for evolutionists, for he believes that punctuationism gives evolutionists the ability to counter creationist arguments. Creationists, Stanley believes, argued against gradualism, and now that punctuationists can explain why gradualism is not the best way to explain evolutionary change, they have an answer for why the fossil record does not validate gradualism. Stanley also asks if the acceptance of punctuational model would throw the whole theory into doubt. He believes this will not happen because although evolutionists debate the patterns and the mechanisms of evolution, they agree that evolution happened. (pg. 166)
Stanley believes that creationism is religion, and not science, in part because of its religious homogeneity and lack of a presence outside of the United States. He resists public debates with creationists because it turns science into something more like a popularity test. This poses a problem for evolutionists, because presenting the case for evolution cannot be done in a few minutes, but might require an entire a college course. (pg. 177) Stanley concludes this chapter predicting that genetics and embryology will continue to show how quantum speciation is possible, strengthening the punctuational model. (pg. 179)
Another danger Stanley perceives to science is that of teleology and orthogenesisthe view that humans are the pinnacle of evolutionary change, and that we were destined to exist through an evolutionary process. Orthogenesis is inappropriate for science because even the Modern Synthesis implied that variation was random, and environmentally-caused selection pressures are impossible to predict. Evolution is thus neither progressive nor regressive.
Stanley believes that the punctuational view only increases the random nature of evolution because it is essentially impossible to predict the direction that speciation will take a species. (pg. 181) Speciation is like experimentation where populations experience temporary chance factors as they probe out the environment to see what works. (pg. 181) Evolutionary trends are possible, where species will over time tend to go in one direction. Yet this is most likely the result of the ability to undergo speciation. Two factors control the ability of a species to undergo speciation: 1) Long-lived species tend to bud off more descendant species. 2) Differential fecundity (i.e. fecundity in terms of the ability of a species to undergo speciation and produce new species) Thus, according to Stanley, just like how individuals in a population which leave the most descendants tend to dominate the future population, so do species which tend to speciate off the most new species, tend to dominate the biota of the future. This is called species selection. Because natural selection within a species does not necessarily affect what goes on when new species are produced, and because speciation is highly unpredictable, microevolution is essentially decoupled from macroevolution. Furthermore, traditionally important questions like What drove humans down from the trees? may become less important under the punctuational view, for major changes can happen in the absence of long-term environmental trends. (pg. 186) In other words, Stanley might say that long-standing environmental pressures did not drive them out of the trees, it just happened as a part of an unpredictable event during speciation.
Species selection can be influenced by environmental trends, but macroevolutionary trends in a direction are most likely to occur simply because a particular type of species tends to be long-lived, or happens to rapidly bud off descendant species. When higher taxa go extinct, it is probably because the species within that taxa tend to not have the qualities enabling them to succeed in species selection. It is possible that species need not be long-lived to be successful. For example, cichlids may have many species purely due to their virtue of having the ability to speciate rapidly (i.e. their populations can quickly establish reproductive isolation). Some plant species also exhibit this quality, due to their ability to easily achieve reproductive isolation due to pollination mechanisms. Thus, macroevolution takes place in response to environmental changes only to the extent that species within a taxa find a way to become long-lived and leave many descendant species.
Extinction also plays a role in macroevolution. Extinction has little to do with microevolution because natural selection takes place when some members of a population reproduce better than others, whereas extinction is when all members of a population fail to reproduce. At this point, Stanley believes that emergent properties arise making macroevolution irreducible to microevolution. (pg. 184-185) Yet, mass extinction does allow the environment to have large effects upon macroevolution, as environmental changes will often select out many types of species.
Many fauna today lack plenitude, or the full range of possible diversity. Though we have large mammals today, such as the elephant, rhino, and hippo, in the not-so-distant-past many more species of large mammals existed. Similarly, before the ice age, there were many more species of shelled mollusks in the Caribbean than there are today. Some extinctions can occur simply because one predator hunts its prey to death, at least in a local region. Predation can eliminate many members of an otherwise very-successful species, thus preventing that otherwise successful species from monopolizing the habitat. Yet when that predator is removed, that successful prey-species can take-over. Ironically, predation can actually allow for higher species diversity by keeping some species in check, allowing more room for others.
Darwin didnt recognize that predation can allow for an increase in species diversity. Darwin thought that there was always this fierce competition in nature for resources, and did not realize that it is possible for many species to coexist. Stanley believes that when predation keeps population numbers of many species in check, competition for living space is not a major environmental factor affecting the success of a species. However, Stanley does believe that this competition might have been a factor in the Pre-Cambrian environment before there were multicellular species, and there likely were not widespread predators floating around keeping the algae and bacteria in check. Plenitude was possible in this environment where the ecospace was saturated with all the primitive autotrophs it could handle. Stanley believes that widespread bacterial mat stromatolite fossils from the PreCambrian indicates that plenitude existed in this environment. Once consumers (i.e. predators) arose, stromatolites began to decline. Stanley thus finds it remarkable that for much of Earths history, plenitude may have reigned, even though today he believes it does not.
Interestingly, Stanley provides a rationale for why the existence of plenitude caused it to take so long for multicellular creatures to arise. With plenitude, the ecosystems were saturated with organisms, and it was difficult for reproductive isolation (and thus speciation) to occur. New breeds of consumers could not arise without the possibility of speciation. Stanley envisions that occasional speciation events may have eventually created the multicellular creatures which started eating the one-celled world of algae and bacteria. Stanley believes that under the punctuational model, it was inevitable that the origin of animals would be followed by an adaptive radiation.
Stanley also interprets the origin of sexual reproduction under a punctuational model. Sexual reproduction entails genetic recombination, and this can allow for large phenotypic changes. The punctuational model might explain the prevalence of sex, because it fundamentally allows for more speciation. (pg. 198) Without sex, the only way to get genetic diversity is to allow for the slow accumulation of mutations, thus slowing diversification. Stanley believes this is why today, sexual creatures are more prolific they simply are better able to produce the variability which aids speciation. Thus, when the environment changes, asexually reproducing creatures are subject to extinction if they cant produce the variability which will allow them to survive. Stanley says that perhaps the only highly successful asexual creature is the bdelloid rotifer, which requires very little diversification to avoid extinction because they can hibernate in the absence of hospitable environmental conditions.
Conclusion I hope this summary accurately represents Stanley's views. I realize that there are many differing views of the fossil record out there, from ID proponents to creationists, and even within the evolutionist community. Whatever you believe, my only hope is that you will learn something about Dr. Steven Stanley's own views and about the punctuational model of evolution by reading this summary. I also hope that if you disagree with anything written here, it is Stanley's own views with which you disagree with and not any misrepresentations of his views which have unintentionally crept into this summary. I thank you for reading and I hope that you find this summary useful, as it was enjoyable and educational for me to write. I also appreciate comments, suggestions, or any form of constructive criticism which you can send to me at casey@ideacenter.org. [1] Other authors have often referred to the theory as punctuated equilibrium or punctuated equilibria however because Stanley refers to it as the punctuational model, the punctuational view, or punctuationalism, those phrases will be used primarily in this book review. [2] Please feel free to send any feedback, comments, or criticisms as I appreciate such and take them seriously. The author may be contacted at casey@ideacenter.org. [3] Of course many have argued that in a broader sense, these random processes are overall statistically predictable such that over time they are not truly random. Nonetheless, Stanley here describes them as random. [4] The first four chapters of TNET tell a fascinating history of evolutionary theory. But all histories must be read in the context of their historian. I do not mean to question Stanley's account, but readers should keep in mind that the history given by Stanley is rooted in his own viewpoint as an evolutionist paleontologist who often questions the importance of the gradualistic model. [5] Evaluating the strength of these dysteleological arguments is beyond the scope of this summary. [6] From what Stanley says, it seems to me that Darwin showed a great measure of respect to those who dissented from his theory and did not resort to accusations or namecalling. [7] Stanley himself doesnt mention the precise age Kelvin actually proposed for the age of the Earth, but I mention it here for clarity. [8] Stanley also attempts to give an interesting morphological rationale for why rapid diversification took place in cichlids. Many of the species are separated by feeding habits. Studies of cichlids have shown that the rear portion of their jaw is decoupled from the front, making it possible for these fishes allow their front teeth to become specialized for one form of feeding, and their back teeth specialized for grinding the food brought in by the front teeth. Thus, many different forms of feeding can be easily imagined from the cichlid jaw makeup.
[9] It should be noted that Stanleys book was written when regulatory genes were just beginning to be understood, and that it is likely that if he were to write the book today, he would have had a much more extensive discussion on this subject. [10] It should be noted, again, that Stanley wrote in 1981 and some ideas about human evolution have changed since that time. [11] Stanley actually discusses some earlier fossils as well, such as Aegyptopithecus, a 30 million year old primate with apelike teeth, a relatively large brain volume compared to other contemporary fossils, leading Stanley to think it could have been an early ancestor of the apes and the smartest animal in its Oligocene world. (pg. 140) Stanley then recounts that the dryopithecine apes lived in the latter Oligocene, and these did give rise to modern apes. Ramapithecus lived in Africa and Eurasia from 14-8 Ma (million years ago), and had teeth more similar to modern forms. (It should be noted that it is no longer generally believed that Ramapithecus was a direct ancestor to modern humans, or the australopithecines. [12] Homo erectus is also known as Pithecanthropus. [13] By the slender australopithecines, Stanley means Australopithecus afarensis and Australopithecus africanus, which are commonly referred to as gracile. [14] As Stanleys book was written in 1981, young-Earth creationism was far and away almost the only form of creationism which had been popularized at the time. | Learn about Problems with the Punctuated Equilibrium Model |








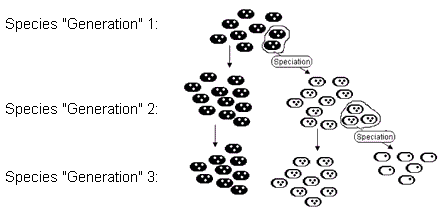
 buds off through a speciation event into species
buds off through a speciation event into species  . Then, after generation 2, species
. Then, after generation 2, species  . In the end, populations representing all three species exist, though all are descended from the original population of
. In the end, populations representing all three species exist, though all are descended from the original population of