Human Origins and Intelligent Design (Less Technical)From The Light Bulb Vol 3:1 (Spring, 2004)by Casey Luskin Editor's Note: This is a less-technical version of an article by the same name that was originally published in the journal Progress in Complexity, Information, and Design in 2005. For the original, more technical version, click here. Two Different Views of Origins
There are two fundamentally different causes for human origins: blind natural processes (chance-law) or purposeful intelligent design. The chance-law hypothesis, neo-Darwinian evolution, states that humans arose through random mutations preserved by blind natural selection. Thus the famous paleontologist George Gaylord Simpson said in his book The Meaning of Evolution, that under evolution, "[m]an is the result of purposeless and natural processes that did not have him in mind."1 On the other hand, intelligent design theory postulates that humans originated due to the intentional arrangement of biomatter--including the human genetic code--by an intelligent agent. Under intelligent design, humans exist because an intelligent being did "have them in mind."
Can we detect intelligent design from the fossil record? One thing we know about intelligent agents is that their complex designs tend to contain large amounts of specified and complex information. Thus, they can rapidly infuse large amounts of genetic information into the biosphere. If this took place in the past, it would be reflected in the fossil record as the abrupt appearance of new fossil forms without similar precursors. Thus, when we find the rapid appearance new fossil forms, which lack transitions from previous different fossil forms, we may infer intelligent design.
Also, designers often re-use basic designs that work, with slight variations. For example, in the parking lot of a mall and youll likely see dozens of cars built upon a similar body design, with slight variations and differences. Similarly, members of a "basic type", represent a group of similar and related species which acquired their genetic programming through intelligent design, and not through common descent.3 Like the car example, members of a basic type are fundamentally similar, but have undergone minor changes through microevolution. These observations can also be built into our understanding of intelligent design.
From our understanding of "basic type" biology, the following could be predictions of intelligent design: 2. Subsequent forms the basic type are variants of, and very similar to, the initial fossil forms.
Some Limitations of Paleoanthropology In 1980 the late paleontologist Stephen Jay Gould noted that, "[m]ost hominid fossils, even though they serve as a basis for endless speculation and elaborate storytelling, are fragments of jaws and scraps of skulls".4 More recently, Nature editor Henry Gee wrote, "[f]ossil evidence of human evolutionary history is fragmentary and open to various interpretations."5 The scarcity of data makes it difficult to confirm how, or even if extinct fossil species are related, and makes it easy to speculate under the influences from preconceptions and biases. Harvard zoologist Richard Lewontin explains: When we consider the remote past, before the origin of the actual species Homo sapiens, we are faced with a fragmentary and disconnected fossil record. Despite the excited and optimistic claims that have been made by some paleontologists, no fossil hominid species can be established as our direct ancestor.6 A Science article entitled, "The Politics of Paleoanthropology," describes how this lack of data causes paleoanthropologists to face challenges in remaining objective because of the sheer lack of evidence and the nature of the subject of study: The field of paleoanthropology naturally excites interest because of our own interest in origins. And, because conclusions of emotional significance to many must be drawn from extremely paltry evidence, it is often difficult to separate the personal from the scientific disputes raging in the field.7 Paleoanthropology is a field where theories may be based only upon limited and incomplete evidence, which is rarely examined through intelligent design.
Many textbooks show interpretive drawings of hominids which may mislead the public to believe actually represent real data (see Figure 2).8 These reconstructions are only loosely based upon fossil evidence and often provide only a highly subjective evolutionary interpretation. As famed physical anthropologist Earnest A. Hooton from Harvard University cautioned in 1931, "alleged restorations of ancient types of man have very little, if any, scientific value and are likely only to mislead the public."9
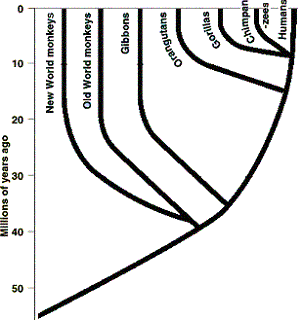
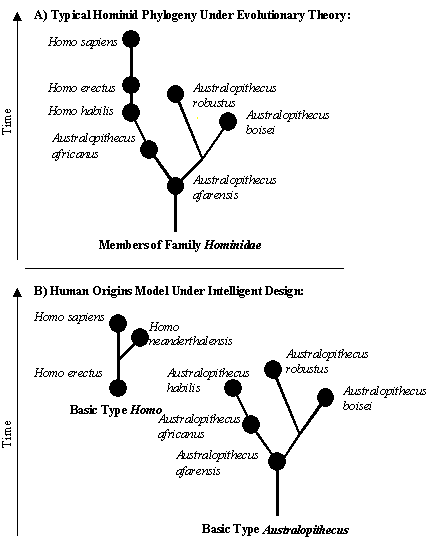
The Australopithecines Humans, apes, and monkeys are members of the Order Primates. Under evolution, all primates are related and the chimpanzee is the closest living relative to humans, and humans are descended from a common ancestor they shared with chimpanzees (see Figure 3). There is essentially no fossil evidence of the supposed evolutionary ancestors of chimpanzees and other living apes,2 however there are some species believed by evolutionists to be ancestors, or close relatives of the ancestors of humans. The majority of "hominid" fossils have been divided into two taxonomic categories: the genus Australopithecus and the genus Homo (which includes our species, Homo sapiens).
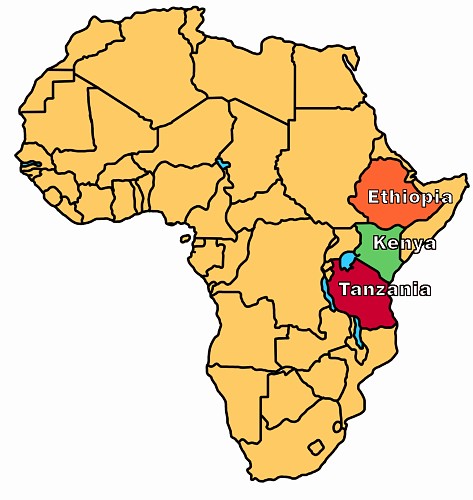
Australopithecines (literally meaning "southern ape") are a genus of extinct hominids that lived in eastern Africa (see Figure 4) from about 4.2 million years ago (Ma) until about 1 Ma.10 Some evolutionists think they are ancestral to humans (see Figure 9), however it has also been argued they are a "side-branch" of the line that led to humans, and not direct human ancestors.12 The four most common species are Australopithecus afarensis, Australopithecus africanus, Australopithecus robustus, and Australopithecus boisei.13 The two smaller and "gracile" forms, africanus and afarensis (the species which includes the famous fossil "Lucy", see Figure 5) are thought by evolutionists to be those most closely related to humans (see Figure 9).
Australopithecines stood about 1-1.5 m in height and had relatively small brains between 370 and 515 cubic cm (cc)14, 15--a range that extends only slightly beyond the brain size of a chimpanzee (see Table 1). Though there are fossils creating a general grade of increasing skull sizes from Australopithecus into modern Homo, the fossil record indicates that about 2 Ma, skull sizes began a "dramatic trajectory" that ultimately resulted in an "approximate doubling in brain size."14 This "rapid evolution" is not uncommon with regards to the origins of characteristics of the genus Homo.
The australopithecine mode of locomotion has been a point of controversy. Many evolutionists believe they were "bipedal" (i.e. walked on two legs). Early studies thought the pelvis of australopithecines was a clear-cut precursor to Homo-like bipedality,16 while many later studies of australopithecine locomotion found it to be different from that of modern apes, but also very different from that of humans--a distinct mode of locomotion.12, 17 One study found sharp differences between the pelvic bones of australopithecines and Homo, and, lacking intermediate fossils, proposed a period of "very rapid evolution corresponding to the emergence of the genus Homo."18 Other recent studies have found that the handbones of Lucy are similar to those of a knucklewalking ape,19, 20 and that their inner ear canals, responsible for balance and related to locomotion, resemble small inner-ear canals of the great apes rather than larger canals found in humans and other members of the genus Homo.21 The most common consensus is that australopithecines were adapted for both tree-climbing and at least semi-upright walking,25 walking differently from humans and living apes.50
However, australopithecines were apes and were very different from humans. One reviewer said that ecologically speaking, australopithecines "may still be considered as apes."23 Harvard paleoanthropologist William Howells mentioned that the arboreal bipedalism of Lucy was "successful in serving Lucy's purposes," but "not something simply transitional"50 to the locomotion of modern humans. These are important clues as to whether or not australopithecines were fully bipedal hominids and ancestral to humans.
It is difficult to connect the australopithecine fossils with any previous fossil primates. Paleoanthropologist Tim White called the early record of hominids "a black hole in the fossil record."22 Paleontologist Steven Stanley notes that, "the latest Miocene and very earliest Pliocene (the period from about eight to four million years ago) has revealed little of the assumed transition to the australopithecines."24 One recent study found that Australopithecus africanus (similar to Australopithecus afarensis), had a body shape more similar to modern apes than to members of Homo.25 Given the distinct qualities of australopithecines and the fact that their skeleton resembled modern apes more than they resemble modern humans,25 it does not seem unreasonable to infer that they could be a designed basic type unrelated to Homo. To strengthen this claim, it remains to be seen if there are species linking Australopithecus to Homo from the fossil record, and the nature of fossil record of the genus Homo must be examined.
Is there a Link between Homo and Australopithecus? Similar to Australopithecus, the genus Homo has a number of different taxonomic schemes. The three most commonly used species include Homo habilis, Homo erectus, and Homo sapiens, which many evolutionists believe are sequentially related, in that order. But there is disagreement. Some "lump" all Homo fossils under a single species, Homo sapiens, while others employ the aforementioned species and Homo neanderthalensis (also classified under Homo sapiens), Homo heidelbergensis (a variant of Homo neanderthalensis), Homo ergaster (similar to Homo erectus), and Homo rudolfensis (otherwise classified as Homo habilis).
One fossil claimed as an intermediate between Australopithecus and Homo is Homo habilis (see Figure 6).11, 26 Homo habilis remains were first discovered in 1960, and were named in 1964 by famous paleoanthropologist Richard Leakey, and his team.13, 26 Despite controversy over fossil dates, habilis, is also often claimed as the earliest member of Homo. Homo habilis is said to show brain enlargement, the first usage of primitive stone tools, and the origin of a humanlike bipedal pelvic gait.26 While it is generally accepted that Homo habilis is a real species, one paleontologist called it a "wastebasket taxon"27 due to the "motley" bone assortment attributed to it. The most complete specimens are only two very fragmented skeletons.13
A study of one of the fragmented specimens compared the post-cranial skeleton of habilis to that of Lucy, supposedly an australopithecine precursor to Homo habilis.28 The study found that Homo habilis was in 24 out of 28 test characteristics more similar to modern African apes than it was to other members of Homo. Given that the skeleton was placed in the genus Homo, these results were called by one of the authors, "unexpected in view of previous accounts of Homo habilis as a link between australopithecines and humans."29
A similar analysis by paleoanthropologists Bernard Wood and Mark Collard found that H. habilis has body proportions more similar to the australopiths than to Homo.25 The study found that the mode of locomotion of Homo habilis was "terrestrial bipedalism with an ability to climb proficiently," and grouped it with the australopithecines. Homo habilis was found to be australopithecine in all of its major characteristicsbody size, body shape, locomotion, jaws and teeth, development, and brain size. Finally, an analysis of the ear canals, indicative of the mode of locomotion, found that a habilis skull is most similar to cercopithecoids (baboons), suggesting it "relied less on bipedal behaviour than the australopithecines."21 This strengthens the case that Homo habilis is not a species of intermediate morphology between australopithecines and Homo, as it lacks reliable criteria connecting it to modern humans, or establishing it as a link between australopithecines and Homo.
After removing habilis from the genus Homo, the earliest known member of Homo becomes Homo erectus (dated as early as 1.9 Ma25). Even if habilis did bear a close resemblance to the genus Homo, it could not be a transition because it appears about the same time as the earliest members of Homo, and most habilis specimens post-date the appearance of Homo.30
A "Big Bang" origin of Homo One study in the Journal of Molecular Biology and Evolution found that Homo and Australopithecus differ significantly in brain size, dental function, increased cranial buttressing, expanded body height, visual, and respiratory changes (see Figure 7):30, 32 We, like many others, interpret the anatomical evidence to show that early H. sapiens [H. erectus and H. ergaster] was significantly and dramatically different from australopithecines in virtually every element of its skeleton and every remnant of its behavior.30
Noting these many changes, the study called the evolutionary origin of humans, "a real acceleration of evolutionary change from the more slowly changing pace of australopithecine evolution"30 and noted that this transformation must have included radical changes: The anatomy of the earliest H. sapiens [H. erectus and H. ergaster] sample indicates significant modifications of the ancestral genome and is not simply an extension of evolutionary trends in an earlier australopithecine lineage throughout the Pliocene. In fact, its combination of features never appears earlier...30 These rapid, unique, and genetically significant changes are termed "a genetic revolution" where "no australopithecine species is obviously transitional."30 One commentator proposed this evidence implies a "big bang theory" of human evolution.33 Although Hawks et. al.30 explain the rapid origin of Homo as an extreme population bottleneck during a "speciation event," the evidence matches the criteria for inferring intelligent design. Given the lack of fossil forms providing sufficient evidence of an evolutionary transformation between Homo and Australopithecus, and given the apparent very large and rapid genetic changes associated with the origin of Homo, from an intelligent design perspective, the "big bang" origin of Homo represents the exact kind mass-infusion of genetic information into the biosphere that would be expected had the genus Homo been intelligently designed apart from relation to Australopithecus.
Homo as a basic type Intelligent design theory predicts that organisms which are reproductively compatible are said to be members of the same basic type.3 Donald Johanson suggested that Homo erectus could reproduce with modern humans.34 Others have suggested that given range of variation within modern humans, Homo erectus could be a member of our modern species.35 Though the postcranial skeleton of Homo erectus is poorly understood, known remains are consistent with modern human mode of locomotion.25 Homo ergaster, often classified under Homo erectus, was found with a nearly complete skeleton that is very similar to modern humans.25 Homo erectus is the "earliest species to demonstrate the modern human semicircular [ear] canal morphology,"21 previously noted as a feature indicative of the mode of locomotion. Some authors have even referred to Homo erectus and Homo ergaster as "early Homo sapiens."30, 36
Wood and Collard found that Homo ergaster and Homo erectus had intermediate cranial capacities,25 but they are within human variation (see Table 1). Not only is cranial capacity of uncertain importance for determining intelligence,31 but erectus skulls as large as 1250 cc38 are within the "normal" range for humans.11 More importantly, erectus remains have been found with signs of culture39 and contemporary with modern humans.40
Later members of Homo such as Homo neanderthalensis are more humanlike. "Neanderthal" fossils have been called a "race" of our own species41, 42 as their body shapes are within the range of modern human variation, 25 and they probably interbred with modern humans.42 Even some evolutionists have speculated they had normal language capabilities.43 and Neanderthal remains have with been found with art,42 culture,42, 43 including burial of their dead,44 and technology including the usage of complex tools,42, 43 and chain-mail armor.45 These similarities make mating compatibility between Homo erectus, Homo neanderthalensis, with modern humans (Homo sapiens) a strong possibility. Neanderthal and Homo erectus differences from Homo sapiens are small and be explained as microevolutionary effects of "size variation, climatic stress, genetic drift and differential expression of [common] genes."49
Conclusion In conclusion, our genus Homo appears to have been intelligently designed and is not connected to the australopithecine apes or any other apes through ancestry:
References: 1. Simpson, G. G., The Meaning of Evolution, pg. 345 (Yale University Press, 1967). 2. Gee, H., In Search of Deep Time: Beyond the Fossil Record to a New History of life (1999). 3. Scherer, S., "Basic Types of Life" in Mere Creation, edited by Dembski 195-211 (1998). 4. Gould, S. J., The Panda's Thumb, pg. 126 (W.W. Norton & Company, 1980). 5. Gee, H., "Return to the planet of the apes," Nature, 412:131132 (July 12, 2001). 6. Lewontin, R. C., Human Diversity, pg.163 (Scientific American Library, 1995). 7. Holden, C., "The Politics of Paleoanthropology," Science, p.737 (August 14, 1981). 8. There are many examples. For one prime example, see Biology: The Dynamics of Life (Glencoe, McGraw Hill), pgs. 438, 442, 443. 9. Hooton, E. A., Up From The Ape, p. 329 (McMillan, 1931). 10. Leaky, M. G., Feibel, C. S., McDouball, I., Walker, A., "New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya," Nature, 376:565-571 (1995). 11. Campbell, N. A., Reece, J. B., Mitchell, L. G., Biology 5th Edition, pgs. 656-665 (1999). 12. Oxnard, C. E., "The place of the Australopithecines in human evolution: grounds for doubt?," Nature, 258:389-395 (Dec 4, 1975). 13. Walker, A., "The Origin of the Genus Homo," in The Origin and Evolution of Humans and Humanness, edited by D. Tab Rasmussen, pgs. 29-47 (1993). 14. Falk, D., "Hominid Brain Evolution: Looks Can Be Deceiving," Science, 280:1714 ('98). 15. Conroy, G. C., et. al., "Endocranial Capacity in an Early Hominid Cranium from Sterkfontein, South Africa," Science, 280:1730-1731 (1998). 16. See Lovejoy, C. O., Kingsbury G. H., Burstein A. H., "The Gait of Australopithecus," American Journal of Physical Anthropology, 38:757-779 (1973). 17. Oxnard, C., E., "Human Fossils: New Views of Old Bones," The American Biology Teacher, 41(5):264-276 (1979). 18. Marchal, F., "A New Morphometric Analysis of the Hominid Pelvic Bone," Journal of Human Evolution, 38:347-365 (2000). 19. Collard, M., Aiello, L. C., "From forelimbs to two legs," Nature, 404:339-340 (2000). 20. Leakey, R.E. F., "Further Evidence of Lower Pleistocene Hominids from East Rudolf North Kenya," Nature, 231:241-245 (1971). 21. Spoor, F., et. al. "Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion," Nature, 369:645-648 (1994). 22. Gibbons, A., "In Search of the First Hominids," Science, 295:1214-1219 (2002). 23. Andrews, P., "Ecological Apes and Ancestors," Nature, 376:555-556 (1995). 24. Stanley, S., The New Evolutionary Timetable, pg. 141 (1981). 25. Wood, B. and Collard, M., "The Human Genus," Science, 284:65-71 (1999). 26. Wood, B., "Origin and evolution of the genus Homo," Nature, 355:783-790 (1992). 27. Tattersall, I., "The Many Faces of Homo habilis," Evolutionary Anthropology, 1(1):33-37 (1992). 28. Scherer-Hartwig, S., and Martin, R. D., "Was "Lucy" more human than her "child"? Observations on early hominid postcranial skeletons," Journal of Human Evolution 21:439-49 (1991). 29. Hartwig-Scherer, S., "Apes or Ancestors" in Mere Creation, edited by William Dembski, pgs. 212-235 (1998). 30. John Hawks, Keith Hunley, Sang-Hee Lee, and Milford Wolpoff, Population Bottlenecks and Pleistocene Human Evolution, Journal of Molecular Biology and Evolution, Vol. 17(1):2-22 (2000). 31. Deacon, T. W., "Problems of Ontogeny and Phylogeny in Brain-Size Evolution," International Journal of Primatology, 11(3):237-282 (1990). 32. Wolpoff, M. H., Paleoanthropology 2nd edition, McGraw-Hill, New York (1999). 33. University of Michigan News and Information Services News Release, "New study suggests big bang theory of human evolution" (January 10, 2000), available at http://www.umich.edu/~newsinfo/ Releases/2000/ Jan00/r011000b.html" (accessed 10/21/03) 34. Johanson, D. C., and Edey, M. A., Lucy: The Beginnings of Humankind, pg. 144, 363 (1981). 35. in, W. S., "Eskimos and Aleuts: Their Origins and Evolution," Science, 142:633-645 (1963). 36. Jelinek, J., "Homo erectus or Homo sapiens?" Rec. Adv. Primatol. 3:419429 (1978). 37. Molnar, S., Races, types, and ethnic groups: the problem of human variation, pg. 57 (1975). 38. Molnar, S., Human Variation Races Types and Ethnic Groups, pg. 65 (2nd ed., 1983). 39. Von Koenigswald, G. H.R., Meeting Prehistoric Man, pgs. 65-79 (1956). 40. Swisher, C. et. al. "Latest Homo erectus of Java: Potential Contemporaneity with Homo sapiens in Southeast Asia," Science, 274:1870-1874 (1996). 41. Delson, E., "One skull does not a species make," Nature, 389:445-446 (1997). 42. Wong, K., "Who were the Neandertals," Scientific American, Aug 25, 2003, pg. 28-37. 43. Textbook: Essentials of Physical Anthropology, 2004, (Thomson Wadsworth). 44. Trinkaus, E., Shipman, P., "Neandertals: images of ourselves," Evolutionary Anthropology, 1(6):194 - 201 (1993). 45. Notes in Nature, 77:587 (April 23, 1908). 46. The Evolution Store, Teaching Quality Fossil Hominids: Reconstruction of Homo erectus 'Peking Man' Replica. http://www.evolutionnyc.com/ IBS/SimpleCat/Product/asp/product-id/488185.html 47. The Evolution Store, Teaching Quality Fossil Hominids: http://www.evolutionnyc.com/IBS/SimpleCat /Product/asp/product-id/488177.html 48. University of Glasgow Hunterian Museum and Art. http://www.hmag.gla.ac.uk/ Neil/labwork/Museumlab.htm 49. Hartwig-Scherer, S., "Apes or Ancestors" in Mere Creation, edited by Dembski, pgs. 212-235 (1998). 50. Howells, W., Getting Here The Story of Human Evolution, pg. 69-85 (1993). |
| ||||||||||||||||||||||||||||||||||||||||